Сочленение - кольцо
Cтраница 1
Сочленение колец у сапсгенинов в точности такое же, как и у стериков. [1]
Сочленение колец А / В и наличие в них двойной связи может влиять не только на выход, но и на направление гидроксилирования. Так, 5р - изо-мер прегнанол - Зр-она - 20 гидроксилируется Rhizopus arrhizus и R. Следует напомнить, что оба организма гидроксилируют прогестерон ( IV) в 6р - и 1 la - положения. Таким образом, наблюдается параллелизм в гидроксилировании А4 - 3-кетостероидов в бр-положение, а их 4 5-дигидропроизводных и Д5 - Зр-оксианалогов - в 7р - положение. Более того, все организмы, способные проводить 7р - гидроксилирование, спо собны также и к Gp-гидроксилированию. Это делает весьма вероятным предположение, что образование 6р - и 7р - оксипроизводных осуществляется одной и той же ферментной системой, которая атакует разные положения субстрата в зависимости от его строения. [2]
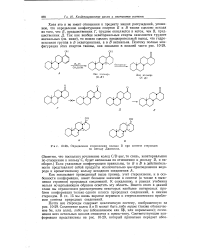 |
Определение стереохимии кольца D при синтезе стероидов. [3] |
Сочленение колец А и В может быть либо транс ( также обозначаемое 5а, или алло), либо цис ( обозначаемое как 5р или нормальное); сочленения всех остальных циклов относятся к транс-типу. [4]
 |
Классификация стероидов по величине углеводородного радикала R у С-17. [5] |
Сочленение колец А и В, В и С, С и D может быть цис - или транс -, как в цис - и граис-декалинах. [6]
Сочленения колец А-В и С - - D у всех буфоге-нинов и близких к ним соединений осуществлены так же, как и у сердечных глюкозидов, рассмотренных выше. [7]
Сочленение колец D и Е - по соседству с гидроксильной группой также должно быть аксиальным. Та - ким образом, образование лактона приводит к конверсии конформации кресла и обращает экваториальную и аксиальную природу эпоксидных связей. [8]
Экваториально-аксиальное сочленение колец п нс-декалпне допускает значительную гибкость системы п здесь снова, как в цнкло-14 ксане, возможна полная конверсия молекулы, идущая через промежуточные ваннообразные формы. Подобное превращение допускает экваториально-аксиальные переходы л гобой группы в любом положении, п, следовательно, заместитель в любом положении п пространственном отношении к месту сочленения колец может быть экваториален. Отсюда следует, что конформацнопный анализ в том виде, is каком он применяется для жестких систем, не может в случае н нс-декалннов принести к определенным выводам о стереохимии молекулы. Для нсех стереонзомерон конформации, в которых гидроксил экваториален ( XLПа, ХЫПб), имеют почти одинаковую свободную энергию. [9]
Сочленение колец Аи В следует из гладко протекающего сужения кольца А различных Зр-оксипроизводных, катализируемого пятихлористым фосфором ( см. разд. [10]
Сочленение колец D и Е вызывает значительное экранирование Д12 - связи ( см. разд. Другие различия в химии а - и 3-амирина, зависящие от наличия в первом из них экваториальных 19 ( 3 - и 20а - метильных групп, были уже обсуждены ( см. разд. [11]
Сочленения колец А-В и С-D у всех буфоге-нинов и близких к ним соединений осуществлены так же, как и у сердечных глюкозидов, рассмотренных выше. [12]
Сочленение колец обойм подшипника с валом и корпусом производится по одной из неподвижных посадок ( Гр - горячая, Пр - прессовая, Г - глухая, Т - тугая и др.) с соблюдением установленных допусков. Подготовленный подшипник в течение 15 - 20 мин. Нагретый подшипник быстро устанавливают и легкими ударами или нажимом добиваются посадки подшипника на место. Посадку подшипника выполняют с помощью различных прессов и оправок. [13]
 |
Винтовые съемники, применяемые при демонтаже подшипников качения. [14] |
Сочленение колец обойм подшипника с валом и корпусом производится по одной из неподвижных посадок ( Гр - горячая, Пр - прессовая, Г - глухая, Т - тугая и др.) с соблюдением установленных допусков. Нагретый подшипник быстро устанавливают и легкими ударами или нажимом добиваются посадки подшипника на место. [15]
Страницы: 1 2 3 4