Схизонты
Cтраница 1
Схизонты циркулируют в гемолимфе хозяина, проникают в ткани, химический состав которых отвечает их требованиям, где оседают и растут в цитоплазме клеток, пока не образуются сферические или яйцевидные, многоядерные схизонты, которые затем вновь делятся на одноядерные меронты. В то время как ядра первых схизонтов в большинстве случаев плотные и равномерно заполнены хроматином, после распада схизонтов появляются ме-розоиты с пузыревидным ядром, с кариосомой в центре. Плазма меронтов вакуолирована и слабее окрашивается красителями. Меронты вновь вырастают в яйцевидные, округлые или лентообразные схизонты, которые вновь делятся на небольшие отрезки. [1]
Червеобразные схизонты разрастаются в грушевидные, позднее в широкояйцевидные схизонты, ядра которых делятся, образуя большое количество дочерних ядер. Вокруг этих ядер формируются и дифференцируются сигаровидные мерозоиты с ядром посредине. Оболочка схизонта лопается, и освободившиеся мерозоиты переходят в ткани, по которым они медленно расселяются вокруг. Во вновь пораженных клетках мерозоиты утолщаются, приобретая ладьевидную, яйцевидную, а затем шаровидную форму. Деление вновь повторяется в зависимости от пространства, которым располагает паразит в жировом теле хозяина. При последующем делении возникают мерозоиты, которые, вырастая, преобразуются в половые стадии, шаровидные гаметоциты. Часть из них остается неизменными, а лишь редуцирует хроматин ядра и таким способом образует гаплоидные макрогаметы, ядро других делится на 64 - 128 дочерних ядер. Макрогамету оплодотворяет только одна микрогамета, хотя в некоторых случаях на макрогамету стремятся осесть несколько микрогамет. После разделения шаровидных споробластов в центре цисты ( зиготы) остается бесформенное остаточное тело. В каждой споре зародыш делится и образуются два спорозоита, изогнутые кольцом вокруг шаровидного остаточного тела. [2]
Трофозоиты и схизонты без псевдоподий. [3]
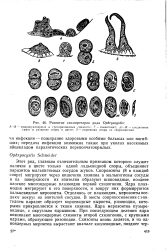 |
Развитие схизогрегарин рода Ophryocystis. [4] |
А-В - микронуклеарные и грегариноидные схизонты; Г - гаеметоцит; Д - Ж - соединение гамет и развитие споры в цисте; 3 - созревшая спора со спорозоитами. [5]
 |
Ускорение процесса отравления жукрв Otiorrhynchus ligustici при пораже - - нии их Nosema otiorrhynchi. [6] |
Вместе со спорами находятся схизонты, способные к дальнейшему делению, как только они окажутся на подходящих для дальнейшего развития тканях. [7]
При заражении мышечных тканей кишечника схизонты проникают между эпителиальными клетками к слою мышечных волокон и оседают в связках непосредственно возле ядер. [8]
Червеобразные схизонты разрастаются в грушевидные, позднее в широкояйцевидные схизонты, ядра которых делятся, образуя большое количество дочерних ядер. Вокруг этих ядер формируются и дифференцируются сигаровидные мерозоиты с ядром посредине. Оболочка схизонта лопается, и освободившиеся мерозоиты переходят в ткани, по которым они медленно расселяются вокруг. Во вновь пораженных клетках мерозоиты утолщаются, приобретая ладьевидную, яйцевидную, а затем шаровидную форму. Деление вновь повторяется в зависимости от пространства, которым располагает паразит в жировом теле хозяина. При последующем делении возникают мерозоиты, которые, вырастая, преобразуются в половые стадии, шаровидные гаметоциты. Часть из них остается неизменными, а лишь редуцирует хроматин ядра и таким способом образует гаплоидные макрогаметы, ядро других делится на 64 - 128 дочерних ядер. Макрогамету оплодотворяет только одна микрогамета, хотя в некоторых случаях на макрогамету стремятся осесть несколько микрогамет. После разделения шаровидных споробластов в центре цисты ( зиготы) остается бесформенное остаточное тело. В каждой споре зародыш делится и образуются два спорозоита, изогнутые кольцом вокруг шаровидного остаточного тела. [9]
В кишечном эпителии этих скорпионов в шаровидных очажках развиваются яйцевидные схизонты и шаровидные цисты, заполненные споробласта-ми. Диаметр цист 34 - 37 мк, веретеновидные споры в них 20 мк длины и 10 мк ширины. Внутри споры имеется всего один спорозо-ит, свернутый в кольцо вокруг небольшого остаточного тела. [10]
Наряду с одноядерными яйцевидными схизонтами в теле насекомого появляются и многоядерные схизонты, которые распадаются на множество меронтов, собранных в группы в форме ягоды шелковицы или малины. Цисты, образовавшиеся в клетках эпителия, где протекает схизогония, разрываются, и меронты выпадают в полость кишечника. С перемещающейся пищей они попадают на непораженные клетки, проникают в них, и там повторяется схизогония. Стадия развития, которая представляла бы гаметы, не обнаружена. В клетках пораженного кишечного эпителия можно обнаружить сферические плазмодии с большим числом ядер, заключенных в прочную пленку и делящихся на 20 - 30 споробластов, шаровидных клеток с одним ядром, которые также заключены в прочную оболочку. Таким путем возникают панспо-робласты, или шаровидные цисты, диаметром обычно 8 мк, в которых относительно свободно лежат шаровидные споры диаметром 1 - 1 5 мк. Цисты из разрушенного эпителия выпадают в кишечник короеда и с экскрементами выходят наружу, деформируясь при этом и приобретая неправильно колоколовидную форму. Цисты очень плохо окрашиваются анилиновыми красителями. [11]
 |
Цикл развития Mycetosporldium jacksonae. [12] |
Наиболее раиней из обнаруженных стадией развития паразита являются шаровидные или яйцевидные одноядерные схизонты, проникающие в клетки поражаемой ткани. Они имеют относительно крупное ядро с округлым хроматиновым зерном в центре. Иногда они вырастают в бесформенные образования, и в них происходит многократное деление ядра, после чего в некоторых случаях вновь отделяются одноядерные меронты, в иных случаях образуются плазмодии, содержащие более 8 ядер. При просмотре некоторых препаратов создается впечатление, что плазмодии имеют ризоидные отростки, что, однако, может быть вызвано препарированием свежего материала. Признаки митотических фигур видны только на некоторых стадиях развития гриба. [13]
Споры открываются в кишечнике гусениц, зародыши проникают в стенки кишечника и превращаются в сферические схизонты размером 0 5 - 1 мк. Через 12 - 48 часов после введения спор в кишечнике гусениц можно обнаружить планонты, однако выбрасывание спорами стрекательных нитей происходит немедленно после прохождения спор вместе с пищей за передний отдел кишечника. Планонты, обнаруживаемые в полости кишечника, происходят, по-видимому, от спор, зародыши которых при выбрасывании стрекательной нити не попали непосредственно на эпителий кишечника. Схизонты, попавшие в гемолимфу - двуядерные. Током гемолимфы они разносятся по всему телу гусеницы и проникают в ткани, где вырастают в шаровидные четырехядерные образования. После деления на одноядерные мерозоиты схизонты начинают делиться дальше, и возникают лентовидные стадии развития длиной до 10 мк, содержащие более 8 ядер. [14]
Схизонты циркулируют в гемолимфе хозяина, проникают в ткани, химический состав которых отвечает их требованиям, где оседают и растут в цитоплазме клеток, пока не образуются сферические или яйцевидные, многоядерные схизонты, которые затем вновь делятся на одноядерные меронты. В то время как ядра первых схизонтов в большинстве случаев плотные и равномерно заполнены хроматином, после распада схизонтов появляются ме-розоиты с пузыревидным ядром, с кариосомой в центре. Плазма меронтов вакуолирована и слабее окрашивается красителями. Меронты вновь вырастают в яйцевидные, округлые или лентообразные схизонты, которые вновь делятся на небольшие отрезки. [15]
Страницы: 1 2 3 4