Телофаза
Cтраница 3
Первая метафаза; десять пар хромосом образуют экваториальную пластинку, которая здесь видна сбоку. Первая анафаза; 10 пар хромосом отходят к разным полюсам. Первая телофаза; хромосомы сливаются в бесформенную массу. [31]
Большинство компонентов матрикса клеточной стенки транспортируется в пузырьках аппарата Голыши к плазматической мембране, где затем выводится из клетга. Химический состав и структура стенки в разных зонах клеточной поверхности различны, поэтому пузырьки с нужными материалами должны избирательно направляться к определенным участкам плазматической мембраны. Эту направленность обеспечивают ( по крайней мере частично) элементы цитоскелета; одним из примеров может служить образование de novo первичной клеточной стенки после митоза ( подробности см. в гл. В конце телофазы между двумя дочерними ядрами остается пучок микротрубочек, расположенных параллельно оси веретена. Этот пучок состоит из двух наборов полюсных микротрубочек веретена, обладающих противоположной полярностью; концы микротрубочек, принадлежащих к разным наборам, перекрываются в дискообразной области, называемой фрагмопластом и находящейся в плоскости экватора бывшего веретена деления. [33]
 |
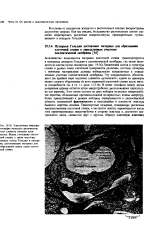
Митоз. На рисунке показана схема митотического деления клетки с одной гомологичной парой хромосом ( Mazia D., Sci. Am., 205, 101 - 120, Sept. 1961. [34] |
В метафазе хромосомы выстраиваются в центре клетки, образуя метафазную пластину, после чего центромера разделяется, и сестринские хроматиды в результате полностью отделяются друг от друга. В анафазе разделенные хроматиды, которые теперь уже называются дочерними хромосомами, движутся к противоположным полюсам так, как будто бы они растаскиваются в результате сокращения волокон веретена. Механизм, лежащий в основе перемещения хромосом, еще не раскрыт. В последней фазе деления клетки - телофазе, во время которой вокруг каждого набора дочерних хромосом образуются новые ядерные оболочки, клетка либо делится на две, либо ( в случае растений) в центре клетки образуются новые плазматические мембраны и клеточная стенка. [35]
Нецеллюлозные компоненты оболочки в силу своей аморфности не имеют определенной ультраструктуры. Оболочки обрабатывают щелочным раствором гидроксиламина, который в этих условиях реагирует с метиловыми эфирами с образованием гидроксамовых кислот. На фото 35 представлена клетка в телофазе митоза; видно, что клеточная пластинка, не достигшая еще продольной стенки, интенсивно окрашена. На этом основании можно сделать вывод, что клеточная пластинка богата метилуронидами. [36]
Первая анафаза постепенно переходит в первую телофазу ( фиг. При этом хромосомы теряют свою четкую форму, слипаются вместе, деспирализуются, вбирают воду и становятся все менее и менее видными. Таким образом достигается стадия покоя, называемая интеркинезом, пли интерфазой ( фиг. Интерфаза характеризуется наличием двух ядер, образованных теми группами хромосом, которые разошлись в анафазе и телофазе. [37]
Ядра размножаются либо прямым делением - амитозом, либо косвенным - кариокинетическим путем, иначе митозом. Ми-тотическое деление ядра наблюдается тогда, когда за ним следует деление клетки. При митозе ( кариокинезе) хромосомы расщепляются вдоль, и число их накануне деления клетки удваивается. В митозе различают четыре фазы: 1) профазу; 2) мета-фазу; 3) анафазу и 4) телофазу. [38]
Продолжительность задержки зависит от кумулятивной дозы в этот момент. Если облучение произведено несколько раньше, кумулятивная доза к моменту наступления критической стадии успевает снизиться, и, следовательно, эффект оказывается меньшим, чем если бы та же доза была сообщена клетке в момент, более близкий к критической стадии. По данным Карлсона, эта критическая стадия, во время которой эффект данной дозы оказывается максимальным и около которой останавливается митотический процесс при задержке деления клеток, приходится на профазу. Это различие, вероятно, несущественно и лишь отражает различие между разными объектами в отношении легкости, с какой можно отличить раннюю профазу от стадии покоя. Так, на нейробластах кузнечика Карлсон ( 1941Ь) показал, что стадия, описываемая как профаза, продолжается вдвое дольше метафазы, телофазы и анафазы вместе взятых и фактически занимает половину всего промежутка времени между телофазой одного деления и метафазой следующего. В тканях же млекопитающих ( Тэнсли, Спир и Глюксман, 1937) 1 и цыпленка ( Юул и Кемп, 1933; Ласницкий, 1940) продолжительность стадии, описываемой как профаза, равна примерно 1 / 3 продолжительности метафазы, анафазы и телофазы вместе взятых. [39]
Продолжительность задержки зависит от кумулятивной дозы в этот момент. Если облучение произведено несколько раньше, кумулятивная доза к моменту наступления критической стадии успевает снизиться, и, следовательно, эффект оказывается меньшим, чем если бы та же доза была сообщена клетке в момент, более близкий к критической стадии. По данным Карлсона, эта критическая стадия, во время которой эффект данной дозы оказывается максимальным и около которой останавливается митотический процесс при задержке деления клеток, приходится на профазу. Это различие, вероятно, несущественно и лишь отражает различие между разными объектами в отношении легкости, с какой можно отличить раннюю профазу от стадии покоя. Так, на нейробластах кузнечика Карлсон ( 1941Ь) показал, что стадия, описываемая как профаза, продолжается вдвое дольше метафазы, телофазы и анафазы вместе взятых и фактически занимает половину всего промежутка времени между телофазой одного деления и метафазой следующего. В тканях же млекопитающих ( Тэнсли, Спир и Глюксман, 1937) 1 и цыпленка ( Юул и Кемп, 1933; Ласницкий, 1940) продолжительность стадии, описываемой как профаза, равна примерно 1 / 3 продолжительности метафазы, анафазы и телофазы вместе взятых. [40]
Основные стадии митоза изображены на фиг. В профазе молекулы ДНК ( ген) реплицируются, хромосома утолщается и удваивается; при этом хромосомы отчетливо вырисовываются в виде двойных нитевидных тел, удерживаемых вместе центромерами. В поздней профазе ядерная оболочка разрушается, а между центросомами образуется веретено. В метафазе неразделившиеся центромеры все еще удерживают вместе хромосомы, однако теперь хромосомы расположены по экватору веретена. В анафазе центромеры разделяются и отходят к противоположным полюсам, увлекая за собой и дочерние хромосомы. В телофазе начинается деление клетки. Намечается образование клеточных мембран. После телофазы наступает новая интерфаза - образуются две дочерние клетки с полным набором хромосом ( и генов), которые идентичны между собой и с хромосомами ( и генами) родительской клетки. В клетке способны к репликации, по-видимому, только хромосомы. Поскольку в репликации зиготы участвуют гены обоих родителей, становится понятным, каким образом достигается строгая передача единиц наследственности от одного поколения другому. [41]
Продолжительность задержки зависит от кумулятивной дозы в этот момент. Если облучение произведено несколько раньше, кумулятивная доза к моменту наступления критической стадии успевает снизиться, и, следовательно, эффект оказывается меньшим, чем если бы та же доза была сообщена клетке в момент, более близкий к критической стадии. По данным Карлсона, эта критическая стадия, во время которой эффект данной дозы оказывается максимальным и около которой останавливается митотический процесс при задержке деления клеток, приходится на профазу. Это различие, вероятно, несущественно и лишь отражает различие между разными объектами в отношении легкости, с какой можно отличить раннюю профазу от стадии покоя. Так, на нейробластах кузнечика Карлсон ( 1941Ь) показал, что стадия, описываемая как профаза, продолжается вдвое дольше метафазы, телофазы и анафазы вместе взятых и фактически занимает половину всего промежутка времени между телофазой одного деления и метафазой следующего. В тканях же млекопитающих ( Тэнсли, Спир и Глюксман, 1937) 1 и цыпленка ( Юул и Кемп, 1933; Ласницкий, 1940) продолжительность стадии, описываемой как профаза, равна примерно 1 / 3 продолжительности метафазы, анафазы и телофазы вместе взятых. [42]
Расхождение гомологичных хромосом к противоположным полюсам соответствует окончанию мейоза I. Число хромосом уменьшилось вдвое, но они все еще состоят из двух хроматид каждая. Если произошел кроссинговер, то эти хроматиды генетически неидентичны и при втором мейотическом делении должны будут разойтись. Веретена и их нити обычно исчезают. У животных и у некоторых растений хроматиды обычно раскручиваются, на каждом полюсе вновь образуется ядерная оболочка и ядро вступает в интерфазу. Затем происходит дробление ( у животных) или формирование клеточной стенки ( у растений) как при митозе. У многих растений не наблюдается ни телофазы, ни образования клеточной стенки, ни интерфазы, и клетка из анафазы I прямо переходит в профазу второго мейотического деления. [43]
Основные стадии митоза изображены на фиг. В профазе молекулы ДНК ( ген) реплицируются, хромосома утолщается и удваивается; при этом хромосомы отчетливо вырисовываются в виде двойных нитевидных тел, удерживаемых вместе центромерами. В поздней профазе ядерная оболочка разрушается, а между центросомами образуется веретено. В метафазе неразделившиеся центромеры все еще удерживают вместе хромосомы, однако теперь хромосомы расположены по экватору веретена. В анафазе центромеры разделяются и отходят к противоположным полюсам, увлекая за собой и дочерние хромосомы. В телофазе начинается деление клетки. Намечается образование клеточных мембран. После телофазы наступает новая интерфаза - образуются две дочерние клетки с полным набором хромосом ( и генов), которые идентичны между собой и с хромосомами ( и генами) родительской клетки. В клетке способны к репликации, по-видимому, только хромосомы. Поскольку в репликации зиготы участвуют гены обоих родителей, становится понятным, каким образом достигается строгая передача единиц наследственности от одного поколения другому. [44]
Возникновение подобной ориентации хромосом происходит при участии клеточного веретена - своего рода системы лучей, идущих от экватора к двум противоположным полюсам клетки. Это веретено имеет форму двойного конуса, в котором отдельные нити или пучки нитей соединены с центромерами хромосом. Эта стадия называется метафазой. Центромеры разделяются, и каждая хроматида приобретает свою собственную центромеру. Таким образом хроматиды превращаются в независимые хромосомы. В течение анафазы к обоим полюсам движутся совершенно одинаковые наборы хромосом. После того как группы хромосом достигли полюса, наступает стадия телофазы ( фиг. Постепенно границы между отдельными хромосомами стираются, и ядро вновь переходит в стадию покоя ( фиг. К этому моменту и вся клетка разделяется на две дочерние, каждая из которых получает по ядру. [45]
Страницы: 1 2 3 4