Тропомиозин
Cтраница 2
Когда мышца в покое ( расслаблена), тропомиозин блокирует на тонком миофиламенте участки для прикрепления миозиновых головок ( рис. 18.24, А), отключая актин. Ионы кальция, высвобождаясь из саркоплазматического ретикулума, соединяются с тропонином, заставляя его и одновременно тропомиозин, с которым он связан, смещаться. [16]
Самая тяжелая, обозначаемая ТпТ, присоединяется к тропомиозину, меньшая ТпТ - к актину. Наименьшая субъединица ТпС может присоединять кальций. Структура комплексов не установлена, так как они не образуют кристаллов. Располагаются молекулы тропонина по одной на каждой молекуле тропомиозина ближе к одному ее концу. Расстояние между соседними молекулами тропонина равно 400 А. [17]
Эта методика была использована для выделения [ i-лактогло-булина, тропомиозина кролика и яичного альбумина; полученные результаты согласуются с данными динитрофенильного метода Сэнджера. [18]
В тонких нитях наряду с актином содержатся другие белки - тропомиозин и тропонин. [19]
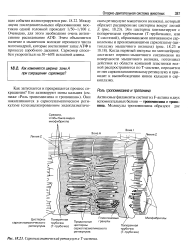 |
Саркоплазматический ретикулум и Т - система. [20] |
Актиновые филаменты состоят из F-актина и двух вспомогательных белков - тропомиозина и тропонина. [21]
В состоянии релаксации ( малая концентрация О2) тропонин, действуя через тропомиозин, каким-то образом подавляет взаимодействие голов миозиновых молекул с актином. При повышении концентрации ионов Са2 1 это препятствие снимается, молекулы АТФ гидролизируются и мышцы сокращаются. Следовательно, комплекс молекул фопонин тропомиозин действует как ингибитор, а ионы Са2 - как активатор мышечного сокращения. [22]
Как отмечалось, кроме рассмотренных основных белков, в миофибрил-лах содержатся также тропомиозин, тропонин и некоторые другие ре-гуляторные белки. [23]
 |
Схема строения тонкой нити. [24] |
В тонких нитях наряду с актином содержатся в меньших количествах другие белки - тропомиозин и тропонин. [25]
 |
Радиационный выход парамагнитных центров при облучении белков и аминокислот. [26] |
Такие белки, как лизоцим, яичный и сывороточный альбумины, пепсин, тропомиозин, содержат различное число глициновых остатков. [27]
Ионы кальция, достигающие при диффузии второго ряда молекул тропонина, с помощью молекул тропомиозина приводят в активное состояние молекулы актина, и происходит гидролиз молекул АТФ на втором ряду голов миозиновых молекул. Энергия гидролиза порождает солитоны в этих молекулах. Их движение внутри толстых нитей вызывает движение новых распухших областей толстых нитей, которые будут продвигать тонкие нити далее к центру саркомера. [28]
Молекулы тропомиозина располагаются вдоль каждой борозды двойной спирали актина, глобула тропв-нина помещается вблизи конца каждой молекулы тропомиозина. [29]
С помощью химических данных, а также результатов рентгено-структурного анализа и электронной микроскопии было показано, что в тропомиозине [213, 223, 224] и в легком меромиозине [ 215J ос-спирали параллельны. По-видимому, это относится и к ос-кера-тину, поскольку длинная цепь а-кератина может быть полностью синтезирована и стабилизирована, прежде чем сможет образоваться суперспираль из антипараллельных а-спиралей. Напротив, в глобулярных белках гемеритрине [216, 217] и оболочке вируса табачной мозаики [180, 218] упаковка спиралей антипараллельна. [30]
Страницы: 1 2 3 4 5