Ключевой фермент
Cтраница 4
Яды смертельны даже в небольших дозах. Известно, что некоторые сильные яды соединяются с ключевыми ферментами, прекращая тем самым катализируемую ими реакцию. Следовательно, в данном случае для проявления действия ядов достаточны лишь небольшие их количества. Для соединения с небольшими количествами фермента нужны небольшие количества яда. Некоторые химиотерапевтические вещества, действующие как антиметаболиты, подавляют ферментные системы микробов. [46]
Механизм действия ДФФ и других нервных ядов показан на фиг. Под влиянием антихолинэстеразной активности нервных газов отравляется лишь один ключевой фермент. Последние составляют активное начало смеси, при помощи которой туземцы Южной Америки отравляют стрелы и дротики. Удаление нервного газа из фермента может быть осуществлено введением противоядия - пиридинальдоксимметиодида. [47]
В свою очередь Са2 1 и цАМФ, активируя ключевые ферменты метаболизма, обеспечивают эффективный ответ клеток на внеш. [48]
 |
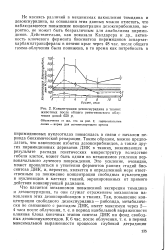
Пути переноса электронов у тионовых бактерий при окислении разных соединений серы. [49] |
При этом бактерии большую часть углерода клеток строят из органических субстратов. Фиксация углекислоты резко снижается из-за подавления синтеза рибулезодифосфаткарбоксилазы - ключевого фермента цикла Кальвина. Снижается также скорость окисления тиосульфата. [50]
 |
Концентрация дезоксиуридина в тканях животных после общего рентгеновского облучения дозой 650 р. [51] |
Не касаясь различий в механизмах накопления тимидина и дезоксиуридина, на основании этих данных можно отметить, что наблюдавшееся повышение концентрации дезоксирибозидов, вероятно, не может быть безразличным для анаболизма пирими-динов. Действительно, как показали Калдарера и др., активность ключевого фермента биосинтеза пиримидинов аспартат-карбамилтрансферазы в печени крыс через 48 час. [52]
Образовавшийся фруктозо-1 6-дифосфат разрывается на две триозы: фосфодиоксиацетон и 3 - ФГА. Разрыв катализируется фрук-тозо-1 6-дифосфатальдолазой ( альдолазой), являющейся ключевым ферментом этого пути. Достаточно обнаружить альдолазу, чтобы получить свидетельство существования гликолитического пути у организма. В последующие реакции может включаться только 3 - ФГА. Фосфодиоксиацетон превращается в 3 - ФГА в реакции изомеризации, катализируемой триозофосфатизомеразой. [53]
Фермент является тетрамером и в тканях млекопитающих представлен тремя изоформами, среди которых наиболее распространены L-пируваткиназа, или печеночный изозим, и Л4 - пируваткиназа, иначе мышечный изозим. Общепризнанным является представление о L-пи-руваткиназе печени как об одном из ключевых ферментов гликолиза. Проблема регуляторной роли Л - пируваткиназы в контролировании гликолиза в скелетной мышце остается нерешенной. Отчасти это связано с тем, что к пируваткиназе мышечной ткани применимы не все критерии, согласно которым, по современным представлениям, фермент может выдвигаться на роль ключевого. В частности, является спорным вопрос об аллостерической природе фермента, а также о проявлении им кооперативных свойств при взаимодействии с субстратами. Возможная причина противоречий связана, по-видимому, с выраженной конформационной подвижностью Mi-изозима пируваткиназы, вследствие чего фермент утрачивает кооперативные свойства при изменении рН или ионного состава среды. [54]
Таким образом, благодаря ресинтезу глутатиона, требующему затраты трех молекул АТФ, цикл может повторяться многократно. Однако это только один из возможных механизмов транспорта аминокислот, поскольку ключевой фермент этого процесса - у-глутамилтрансфераза обладает узкой специфичностью: активен к полярным незаряженным аминокислотам, например цисте-ину, серину; менее активен к дикарбоновым аминокислотам; пролин - вообще таким путем не транспортируется через мембрану. [55]
Многие исследователи считают, что определяющая роль в термофилии принадлежит белкам, в первую очередь ферментным. С этих позиций основные температурные точки термофилов зависят от конформации одного или нескольких ключевых ферментов: при минимальной температуре роста происходит переход от жесткой неактивной конформации белковых молекул к конформации с ограниченной гибкостью; оптимальная температура роста определяет наиболее благоприятное конформационное состояние ферментных белков; при максимальной температуре начинаются нарушения конформации белков и снижение их ферментативной активности, а выше этой температуры рост прекращается вследствие тепловой денатурации белков. [56]
Он также резко увеличивает урожай хлопчатника, сахарной свеклы, томата, редиса. Механизм действия пока не установлен, однако предполагается, что соединение оказывает влияние на ключевые ферменты фотосинтеза. [57]
Страницы: 1 2 3 4