Фотосинтетическое фосфорилирование
Cтраница 2
Как показали сравнительные опыты, при укоренении зеленых черенков в темноте чувствительность фотосинтетического фосфорилирования к ингибиторам в концентрациях, подавляющих рост на 50 %, выраженных в долях от насыщенного раствора, резко падает. [16]
Синтез ( регенерация) АТР осуществляется в основном с помощью трех процессов: фотосинтетического фосфорилирования ( разд. Два первых процесса сходны между собой в том, что АТР образуется в них при участии АТР-синтазы. Субстратное фосфорилирование может происходить при различных реакциях промежуточного метаболизма. В обмене углеводов важнейшие реакции, приводящие к регенерации АТР, катализируются фосфоглщераткиназой, пируваткиназой и аце-таткиназой. Бактерии и дрожжи, сбраживающие сахара, располагают лишь тем АТР, который образуется с помощью этих ферментов. [17]
В растениях витамины группы К участвуют в окислительно-восстановительных процессах и, в частности, в процессе фотосинтетического фосфорилирования, где они служат промежуточными переносчиками электронов. Биосинтез витаминов К происходит в зеленых частях растений и связан с синтезом хлорофилла. [18]
Выдвинуто предположение, что в рекомбинации Н и ОН определенную роль играют цитохромы и что в процессе фотосинтетического фосфорилирования участвует последовательный ряд цитохромов, аналогично тому, как это предполагается для фосфорилирования в дыхательной цепи. При освещении листьев или хлоропластов наблюдаются характерные спектральные изменения. Эти изменения соответствуют возбуждаемой светом реакции окисления цитохрома f гидроксилом и восстановления цитохрома Ьв водородом. Дальнейшим доказательством участия цитохромов служат данные, что циклическое фотосинтетическое фосфорилирование, катализируемое отмытыми хроматофорами Chromatium и JR. Показано, что антимицин А не оказывает значительного подавляющего действия на циклическое фотосинтетическое фосфорилирование в хлоропластах. Однако эти данные необходимо проверить, так как в бактериальных хроматофорах устойчивость к антими-цину А зависит от фактора ( или факторов), содержащегося в над-осадочной жидкости. Гептил-4 - оксихинолин - ] М - оксид является сильным ингибитором нециклического фотосинтетического фосфорилирования в хлоропластах. [19]
АДФ и АТФ занимают центральную роль в энергообмене всех типов клеток, являясь субстратами и продуктами реакций окислительного, субстратного и фотосинтетического фосфорилирования. Энергия, высвобождающаяся при гидролизе АТФ, обеспечивает выполнение всех видов биологической работы. ГТФ энергетически обеспечивает процессы трансляции белков, УТФ необходим для синтеза гликогена, ЦТФ участвует в синтезе глицерофосфолипидов. [20]
В книге Основы фотосинтеза в сжатой и доступной форме излагаются основные вопросы биофизики, биохимии и физиологии фотосинтеза: представления об эволюции фотосинтеза, химическом составе и структуре фотосинтетнческого аппарата, поглощении квантов света пигментами, окислительно-восстановительных реакциях, протекающих с участием хлорофилла, о поглощении и выделении кислорода хлоропластами, фотосинтетическом фосфорилировании и механизме образования АТФ, реакциях усвоения СОз и образовании в процессе фотосинтеза углеводов, органических кислот, аминокислот, о регуляции фотосинтеза, оттоке ассимилятов, связи фотосинтеза и урожая. [21]
Относительные скорости нециклического фотосинтетического фосфорилирования, связанного с восстановлением НАДФ, и циклического фотосинтетического фосфорилирования в целом растении в настоящее время неизвестны. В изолированных хлоропластах скорость фотосинтетического фосфорилирования в присутствии ФМС превышает 2000 мкмоль фосфора на 1 мг хлорофилла в 1 час. Такая скорость более чем достаточна для фотосинтеза, который в неповрежденном листе при полном освещении протекает со скоростью около 200 мкмоль поглощенной углекислоты на 1 мг хлорофилла в 1 час. [22]
Арнон и его сотрудники [71] обнаружили, что освещение растворов хлоропластов в присутствии АДФ и фосфата приводит к образованию молекул АТФ. Этот процесс был назван фотосинтетическим фосфорилированием, в отличие от окислительного фосфорилирования, осуществляемого в митохондриях. В результате многих исследований было установлено, что конечными стабильными продуктами первых световых реакций у высших растений и водорослей являются молекулы АТФ, НАДФ-Ы и 02t у бактерий образуются молекулы АТФ и НАД-И без выделения кислорода. [23]
Подавление реакции фотолиза воды, тормозящей связывание углекислоты и синтез органического вещества в процессе фотосинтеза, наблюдается также при внесении региона и хлор - ИФК. Диурон приводит к депрессии процесс фотосинтетического фосфорилирования. [24]
Добавление к системе каталитических количеств ( приблизительно 3 - 10 5 М) ФМН или витамина Кз ( фиг. НАДФ и вызывает более быстрый циклический тип фотосинтетического фосфорилирования. [25]
Естественно, что в первых опытах интенсивность образования АТФ ( этот процесс получил название фотосинтетического фосфорилирования) была невелика, по сравнению с фотосинтетическим аппаратом in vivo, особенно в опытах с изолированными хлоро-пластами. В дальнейшем интенсивность фотосинтетического фосфорилирования у изолированных хлоро-пластов удалось значительно повысить за счет совершенствования методов их извлечения, применения новых буферных смесей, новых эффективных микрометодов определения количества АТФ. [26]
Следовательно, фотосинтетическое фосфорилирование является тем специфическим источником энергии, который обслуживает процессы роста и органогенеза только на свету. Как в этиолированных тканях отрезков колеоптилеи, так и в зеленых тканях черенков, помещенных в темноту, система фотосинтетического фосфорилирования не функционирует. Окислительное же фосфорилирование функционирует как в темноте, так и на свету одинаково активно. Таким образом, в растягивающихся клетках отрезков колеоптилеи процессы окислительного фосфорилирования доминируют, а синтезы белка и нуклеиновых кислот протекают ослабленно, что и определяет слабую чувствительность отрезков колеоптилеи к ингибиторам нуклеиново-белкового обмена. [27]
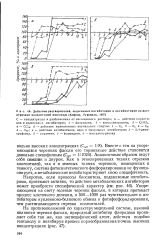 |
Действие растворителей, эндогенных ингибиторов и антибиотиков на рост отрезков колеоптилей пшеницы ( Кефели, Турецкая, 1967. [28] |
Аналогичным образом ведут себя симазин и диурон. Как в этиолированных тканях отрезков колеоптилей, так и в зеленых тканях черенков, помещенных в темноту, система фотосинтетического фосфорилирования не функционирует, а метаболические ингибиторы теряют свою специфичность. [29]

Страницы: 1 2 3 4