Ингибирование - холинэстераза
Cтраница 4
Применение таких соединений в сельском хозяйстве вполне безопасно. Изучение механизма действия органических соединений фосфора на млекопитающих и насекомых показало, что в организме животных они фосфорилируют жизненно важные эстеразы, инги-бируя их нормальные функции. Полагают, что действие этих соединений направляется главным образом на ингибирование холинэстеразы, физиологические функции которой в организм & животных очень важны. [46]
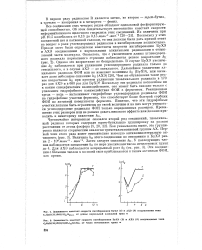
Все соединения этих четырех рядов обладают одинаковой фосфорилирую-щей способностью. Об этом свидетельствует постоянство констант скорости неферментативного щелочного гидролиза этих соединений. Поскольку у этих соединений нет и катионной головки, то они должны были дать прямой ответ на вопрос о роли углеводородных радикалов в ингибировании холинэстераз. Прежде всего были определены константы скорости ингибирования БуХЭ и АХЭ соединениями с нормальными алкильными радикалами в отщепляемой части молекулы. Однако это возрастание не беспредельно. В случае БуХЭ увеличение / сц наблюдается при удлинении углеводородного радикала только до гексильного, а в случае АХЭ - до октильного. Поскольку эти радикалы неспособны ни к каким специфическим взаимодействиям, оно может быть связано только с усилением гидрофобного взаимодействия ФОИ с ферментом. Реакционная среда - вода - выталкивает гидрофобные углеводородные радикалы ФОИ на гидрофобные участки фермента, что способствует более быстрой сорбции ФОИ на активной поверхности фермента. Понятно, что эти гидрофобные участки должны быть ограничены по своей величине И на них могут умещаться углеводородные радикалы ФОИ только определенных размеров. Превышение этих размеров или не должно давать никакого эффекта или должно приводить к некоторому снижению / сп. [48]
На этом основании было высказано предположение, что одним из факторов, определяющих взаимодействие фермента с субстратом при образовании комплекса Михаэлиса, служит ионная реакция между катионным центром ацетилхолина и анионным центром на активной поверхности фермента. Такое взаимодействие должно быть первичным в реакции фермента с субстратом, поскольку ионные силы проявляются на большем расстоянии, чем другие виды химических взаимодействий. Образованию ионной связи приписывали лишь якорную функцию, в результате реализации которой молекула субстрата ориентируется на поверхности фермента, чем значительно облегчается образование других необходимых химических связей ( уже ближнего действия) с группировками активного центра фермента. В связи с этим исследованию кинетики и механизма ингибирования холинэстераз ионами тетраалкиламмония было уделено особое внимание. [49]
Убежденный в правильности этого объяснения, Вильсон сделал далее вывод о том, что при таких обстоятельствах ингибирование можно устранить вытеснением фосфатной группы фермента. Прочная связь между элек-трофильной фосфатной группой и нуклеофильной частью фермента теоретически также должна бы была расщепляться сильно нуклеофильными соединениями. В настоящее время показано, что фермент, инактивированный эфнрами фосфорной кислоты, реактивируется в незначительной степени даже при хранении в воде в течение нескольких недель. Если применяется соединение более нуклеофильное, чем вода, то реактивирование действительно ускоряется. В своих опытах Вильсон применял для ингибирования холинэстеразы ТЭПФ и показал, что гидроксиламин среди реактиваторов действует наиболее быстро. [50]
Не существует четкого разграничения между биомаркерами воздействия и биомаркерами эффекта. Например, можно считать, что образование аддуктов отражает скорее эффект, чем воздействие. Однако биомаркеры эффекта обычно указывают на изменения функции клеток, тканей и всего организма. Некоторые исследователи рассматривают в качестве биомаркеров эффекта такие серьезные изменения как увеличение веса печени у лабораторных животных под воздействием химических веществ и замедленный рост детей. В прикладных целях гигиены труда биомаркеры эффекта следует ограничивать индикаторами субклинических или необратимых биохимических изменений, таких как ингибирова-ние ферментов. Наиболее часто в качестве биомаркера используется, по-видимому, ингибирование холинэстеразы, вызываемое отдельными инсектицидами, такими как фос-форорганические инсектициды и карбаматы. В большинстве случаев данный эффект абсолютно необратим, а ингибирование ферментов отражает общее воздействие данной группы инсектицидов. [51]
Два содержащих фосфаты родственных сложных эфира, крезилдифенилфосфат и о-изопропилфенилдифенилфосфат, также нейротоксичны для некоторых видов животных, включая людей, кур и кошек. Взрослые животные обычно более восприимчивы к нему, чем молодняк. После одной большой дозы этих нейротоксичных органических фосфорных соединений через 8 - 10 дней проявляется поражение аксонов. Хроническое воздействие небольших доз также производит нейротоксический эффект. Механизм воздействия на аксоны периферических нервов, а также на восходящие и нисходящие пути спинного мозга отличен от ингибирования холи-нэстеразы. Поскольку лишь немногие из антихолинэстераз-ных инсектицидов, относящихся к органическим фосфатам, вызывают подобный эффект ( диизопропилфторфосфат, леп-тофос и мипафокс), задержанная невропатия, вероятно, имеет механизм, отличный от ингибирования холинэстеразы. Связь между ингибированием псевдо - или истинной холинэстеразы и нейротоксичным эффектом довольно слаба. [52]
В то же время 0 0-диэтил - 5-зтилтиофосфат ( соединение 1) характеризуется выраженной антихолинэстеразной активностью. Его изомер-триэтилтионфосфат ( соединение 2) - характеризуется в - 20 раз меньшей константой скорости реакции с холинэстеразой и в - 400 раз меньшей константой скорости гидролиза. Это уменьшение реакционноспособности объясняется существенным увеличением энергии активации обоих процессов в случае реакции с ферментом на 5 ккал и для гидролиза - примерно на 6 / с / аи. Любопытно, что предэкспонент при этом даже возрастает приблизительно на 2 порядка, но весьма сильное увеличение энергии активации существенно перекрывает этот эффект. Соответствующий этиловый эфир тионтиолфосфорной кислоты ( соединение 3) занимает по величинам энергии активации и предэкспонента среднее положение. Сопоставление кинетических констант трех соединений - 4 - 6 - подтверждает ту же закономерность. Переход от тиоэфира фосфорной кислоты ( соединение 4) к эфиру тионфосфорной кислоты дает снижение константы скорости ингибирования холинэстеразы на 2 порядка и константы скорости гидролиза - в 20 раз за счет возрастания энергии активации. Этиловый эфир тионтиолфосфорной кислоты здесь также занимает промежуточное положение. [53]
Страницы: 1 2 3 4