Бактериальная хромосома
Cтраница 3
Профаг - генетически активная часть бактериофага, которая в лизогенных бактериях включается в определенный участок бактериальной хромосомы. [31]
Большое значение фактора F для картирования хромосом определяется тем обстоятельством, что изредка он интегрируется с бактериальной хромосомой. С помощью прямой электронной микроскопии было Доказано, что и хромосома, и фактор F имеют кольцевое строение. [32]
 |
Генетическая и физическая карта генома фага А, . Более детальная. [33] |
Блокируя эти операторы, репрессор предотвращает синтез ферментов, необходимых для исключения ДНК фага А, из бактериальной хромосомы, а также для репликации и транскрипции остальных генов. [34]

Транспозоны-это последовательности ДНК, которые способны встраиваться во многие участки генома и могут перепрыгивать с плазмиды на бактериальную хромосому, на другую плазмиду или на умеренный фаг. Транспозоны содержат гены, определяющие внешне распознаваемые признаки, а именно устойчивость к таким антибиотикам, как пенициллин, тетрациклин или канамицин. [36]
В настоящее время известны по меньшей мере три разных механизма рекомбинации попавшей в бактериальную клетку чужеродной ДНК с бактериальной хромосомой ( или с плазмидой) in vivo: 1) общая гомологичная рекомбинация, 2) сайт-специфическая рекомбинация и 3) негомологичная рекомбинация. [37]
Начавшийся процесс репликации хромосомы бактерии продолжается до тех пор, пока не удвоится вся ДНК - В этом смысле бактериальная хромосома представляет собой единицу репликации - репликон. Другие молекулы ДНК, которые могут присутствовать в бактериальных клетках ( см. гл. V), также представляют собой отдельные репликоны. [38]
Начавшийся процесс репликации хромосомы бактерии продолжается до тех пор, пока не удвоится вся ДНК - В этом смысле бактериальная хромосома представляет собой единицу репликации - репликон. Другие молекулы ДНК, которые могут присутствовать в бактериальных клетках ( см. гл. V), также представляют собой, отдельные реп л иконы. [39]
Начавшийся процесс репликации хромосомы бактерии продолжается до тех пор, пока не удвоится вся ДНК - В этом смысле бактериальная хромосома представляет собой единицу репликации - репликон. Другие молекулы ДНК, которые могут присутствовать в бактериальных клетках ( см. гл. V), также представляют собой отдельные репликоны. [40]
Одна бактериальная клетка может одновременно содержать профаги нескольких неродственных умеренных фагов; в этом случае эти профаги включаются в бактериальную хромосому в разных местах. Вместе с тем одна бактерия может содержать лишь одного из мутантов данного штамма умеренного фага. В самом деле, эти мутанты могут включаться лишь в один локус, и если он уже занят, то никакой другой мутант данного штамма не может обосноваться в этой бактерии. Из разных лизогенных штаммов разновидности Escherichia coli К-12 было выделено примерно 30 разных умеренных фагов. [41]
Изучение фага лямбда ( k), лизогенного для Escherichia coli К12, позволило выяснить, каким образом профаг связан с бактериальной хромосомой. [42]
Очевидно, репликация эукариотиче-ской ДНК, которая организована в ну-клеосомы и находится в составе хрома-типовых волокон, должна быть гораздо более сложной, чем репликация бактериальной хромосомы. [43]
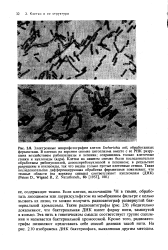
Если клетки, включившие 3Н в тимин, обработать лизоцимом или лаурилсульфатом на мембранном фильтре с целью вызвать их лизис, то можно получить радиоавтограф развернутой бактериальной хромосомы. Такие радиоавтографы ( рис. 2.9) убедительно доказывают, что бактериальная ДНК имеет форму нити, замкнутой в кольцо. Эта нить в генетическом смысле соответствует группе сцепления и называется бактериальной хромосомой. Кроме того, радиоавтографы позволяют представить себе способ деления такой нити. На рис. 2.10 изображена ДНК бактериофага, выявленная другим методом. [45]
Страницы: 1 2 3 4