Белковая цепь
Cтраница 3
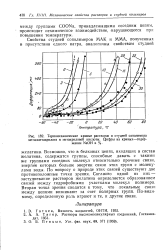 |
Термомеханические кривые растворов и студней сополимера. [31] |
Возможно, что в белковых цепях, входящих в состав желатина, содержатся группы, способные давать с такими же группами соседних молекул относительно прочные связи, энергия которых больше энергии связи этих групп с молекулами воды. По вопросу о природе этих связей существуют две противоположные точки зрения. Согласно одной из них - застудневание растворов желатина определяется образованием связей между гидрофобными участками молекул полимера. Вторая точка зрения ( сводится к тому, что локальные связи между цепями возникают за счет полярных групп. По-видимому, определенную роль играют и те, и другие связи. [32]
Выяснение последовательности аминокислотных остатков в белковой цепи представляет собой очень сложную и трудоемкую задачу, решение которой возможно лишь на основе использования комплекса специальных методов. Прежде всего расщепляют белок на сравнительно короткие фрагменты путем разрыва лишь некоторых амидных связей; для этой цели используют избирательные энзиматические или химические методы. [33]
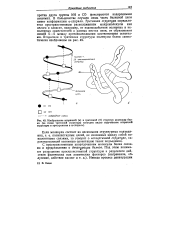 |
Изображение вторичной ( а и третичной ( б структур молекулы белка ( на схеме третичной структуры наглядно видно скручивание вторичной структуры в пространстве в о-спираль. [34] |
В большинстве случаев лишь часть белковой цепи имеет конформацию а-спирали. Третичная структура определяется пространственным расположением макромолекулы как целого и зависит, например, от взаимодействия полярных и неполярных заместителей в разных местах цепи, от образования связей S-S между противолежащими цистеиновыми остатками. [35]
Эта структура состоит из шести белковых цепей, каждая из которых имеет форму а-спирали, закрученной вокруг центральной спиралеобразной белковой молекулы. В результате образуется веревко-образная структура. [36]
В связи с этим простейшая модель белковой цепи использует грубое представление о звеньях двух сортов - гидрофобных и гидрофильных. [37]
Оптическая активность считается необходимым условием при белковой цепи, и, вероятно, без диссимметричных молекул жизнь была бы можна. Следует отметить, что асимметричные молекулы ( обладающие только одним элементом симметрии - осью GI) составляют особую группу диссимметричных молекул, хотя не все диссимме-молекулы являются асимметричными. Стереоизомеры имеют одинаковый скелет, однако различаются абсолютным пространственным рас-атомов. [38]
Задача ставится так: предсказать участки белковой цепи, имеющие определенную вторичную структуру, в данном случае а - спираль. Для предсказания используется только информация о последовательности аминокислотных остатков в молекуле. [39]
Матричная РНК служит шаблоном для сборки белковой цепи. Четырехбуквенный нуклеотидный текст переводится в двадцатибуквенный - аминокислотный. Возникает проблема генетического кода, впервые отчетливо сформулированная физиком Гамовым в 1954 году. [40]
Однако на следующем уровне, в белковой цепи, количество содержащейся информации уменьшится по сравнению с уровнем ДНК, поскольку каждый из 20 аминокислотных остатков кодируется тремя нуклеотидами. [41]
 |
Отнесение аминокислотных остатков к октетам на основании нит. [42] |
Само существование фиксированной первичной структуры у белковой цепи доказывает, что в клетке должна быть заложена программа построения этой структуры. Текст не может возникнуть в результате случайных встреч аминокислот - подобно типографскому тексту он должен набираться на некоторой матрице. [43]
Эти мутации, приводящие к обрыву белковой цепи, особенно опасны. [44]
Вопрос о способе возникновения первичной структуры белковой цепи в процессе матричного синтеза, идущего с необходимым участием других информационных макромолекул ( молекул нуклеиновых кислот), представляет собой сложную физическую проблему. В связи с этим возникает физическая проблема генетического кода. Представление о генетическом коде исходит из предположения о существовании специфического молекулярного механизма превращения генетической информации в структурную функциональность белковых молекул. Это предположение имеет физический характер. [45]
Страницы: 1 2 3 4