Полипептидная цепь - белок
Cтраница 3
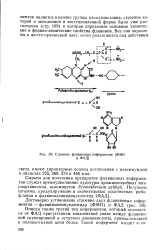 |
Строение флавиновых коферментов ( ФМН и ФАД. [31] |
Описан также третий тип коферментов, который отличается от ФАД присутствием ковалентной связи между флавино-вой группировкой и остатком аминокислоты, принадлежащей к полипептидной цепи белка. [32]
Вскоре после открытия и окончательного признания генетической роли ДНК ( 1944 - 1953) стало ясно, что ДНК не является непосредственной матрицей для синтеза полипептидных цепей белков. [33]
Опыты Левинталя, Герена и Ротмапа дали интересную возможность проверки того, что расстояние между двумя мутантами на генетической карте пропорционально расстоянию между поврежденными точками в полипептидной цепи белка. Поскольку речь идет о мутациях внутри одного цистрона ( конкретно внутри Р - ло-куса), то ясно, что имеются в виду повреждения одного фермента. Этот вопрос является одним из самых фундаментальных в молекулярной биологии. [34]
Природными гликопротеинами являются смешанные биополимеры, в молекулах которых остатки олиго - и полисахаридов ковалентно связаны ( О - или Л - гликозидными связями) с полипептидными цепями белков. [35]
ГЛИКОПРОТЕЙНЫ ( гликопротеиды), соед, в молекулах к-рых остатки олиго - или полисахаридов ковалентно связаны ( О - или М - гликозидными связями) с полипептидными цепями белка. [36]
Он считал, что обнаруженные им закономерности [ описанные уравнениями ( 2, 3, 4) ] основаны на структурных особенностях белков различных классов и, в первую очередь, связаны с последовательностью аминокислотных остатков в полипептидной цепи белка. Этот тезис - аминокислоты не могут располагаться в полипептидной цепи произвольно, их последовательность должна соответствовать найденным закономерностям - ознаменовал появление нового подхода к исследованиям белковых структур. С этого момента у химиков все больше и больше укрепляется представление о том, что ключом к разгадке строения индивидуальных белков является установление последовательности их: аминокислотных остатков. [37]
МАТРИЧНЫЕ РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ ( информационные РНК, мРНК, иРНК), молекулы РНК, представляющие собой комплементарные ( см. Комплементар-ность) копии участков значащих цепей генов ( ДНК), в к-рых закодирована информация об аминокислотных последовательностях полипептидных цепей белков. Образуются в результате транскрипции ( синтез РНК на ДНК-матрице с помощью фермента ДНК-зависимой РНК-полимеразы) и послед, процессинга первичного транскрипта - удаления избыточных РНК-последовательностей с обоих концов молекулы и некодирующих последовательностей РНК, а также сшивания кодирующих участков РНК друг с другом. [38]
Метод Эдмана последовательной деградации пептидов и белков основан на реакции фенилизотиоцианата с концевой аминогруппой в щелочной среде, которая приводит сначала к образованию тиомочевинной метки концевой аминокислоты, последняя при действии CF3COOH отщепляется, в конечном счете, в виде замещенного фенилтиогидантоина, после чего полипептидная цепь белка укорачивается на один аминокислотный остаток. [39]
В случае белков и нуклеиновых кислот важнейшими структурными элементами, изменяющими, по всей вероятности, свои коэффициенты активности в присутствии солей, являются заряженные, гидрофобные или неполярные группы, такие, как алифатические и ароматические боковые цепи аминокислот и основания нуклеиновых кислот, а также полипептидная цепь белков и диэфирная фосфатная цепь полинуклеотидов. [40]
Гидролазы относятся к ферментам, у которых активная группа образована в результате особой конформации белковой молекулы. При изгибах полипептидной цепи белка некоторые аминокислотные остатки сближаются и их сочетание образует активный центр биологического катализатора. Высказывалось мнение, что строгая специфичность фермента ( его узкая специальность - настроенность только на определенную реакцию) зависит от окружения активной группы. [41]
Одним из наиболее исследованных семейств ферментов являются сери-нопротеазы. Все они предназначены для расщепления полипептидных цепей белков по механизму, в котором участвует боковая цепь аминокислоты серина ( - СН2 - ОН), находящейся в активном центре фермента. Три такие протеазы ( трипсин, эластаза и химотрипсин) синтезируются в поджелудочной железе и выделяются ею в кишечник, где они превращают содержащиеся в пище белки в аминокислоты, способные всасываться через стенки кишечника. Благодаря возможности легко изолировать эти ферменты и их сравнительно высокой устойчивости их удалось интенсивно исследовать химическими способами еще до того, как стало возможным проведение рентгеноструктурного анализа белков. В настоящее время биохимический и рентгеноструктурный анализы позволили установить достаточно ясную картину функции этих ферментов, иллюстрирующую два аспекта действия любых ферментов: каталитический механизм и специфичность к субстрату. [42]
После того как аминокислотные остатки расположились на полинуклеотидной цепи РНК в определенной последовательности ( или по мере такого расположения), происходит перегруппировка, заключающаяся в разрыве ангидридных связей и образовании пептидных. Таким образом, на РНК синтезируется полипептидная цепь белка со специфическим порядком аминокислотных остатков, соответствующим определенному строению данной РНК. [43]
Еще на ранней стадии исследования белков было обнаружено, что при некоторых видах их химической модификации, не приводящих к изменениям молекулярного веса, происходит значительное увеличение вязкости растворов белков. Это явление объясняется тем, что полипептидная цепь немодифицированного белка заспирализована и свернута, в результате чего она принимает глобулярную форму. Пример способа сворачивания приведен на рис. 40.5, на котором схематически представлена третичная структура миоглобина, имеющего протяженные спиральные участки, чередующиеся с неспиральной, нерегулярной вторичной структурой; именно на этих участках происходит сворачивание и изгибание. Как было предсказано, а затем подтверждено рентгеноструктурным анализом, пролин вследствие своей циклической структуры образует пептидные связи с углами, исключающими спиральную структуру. [44]
Гипотезу о том, что характер свертывания полипептидной цепи белка ( т.е. его вторичная и третичная структуры) определяются его линейной аминокислотной последовательностью, можно проверить, если дать развернутым белковым молекулам снова самопроизвольно свернуться. [45]
Страницы: 1 2 3 4