Растущая полипептидная цепь
Cтраница 3
Далее следует реакция между ацильной группой дипептида образовавшейся дипептидил - s - PHK и аминогруппой третьей аминокислоты, соединенной с транспортной РНК. Процесс повторяется по мере присоединения каждой следующей аминокислоты к растущей полипептидной цепи. В условиях активного белкового синтеза рибосомы находятся в виде агрегатов, названных полирибосомами ( гл. Полагают, что полирибосомы представляют собой группы рибосом, прикрепленных к одной общей молекуле информационной РНК; рибосомы передвигаются вдоль информационной РНК, причем на каждой рибосоме формируется полипептидная цепь. [31]
Обратное превращение таутомера ( 1106) в азлактон происходит нестереоселектив-но и приводит к рацемизации аминокислотного остатка. Это особенно опасно при твердофазном синтезе, так как в этом случае не используются промежуточные стадии очистки и происходит постоянное накопление диастереомеров в растущей полипептидной цепи. Не существует способов, позволяющих целиком избежать рацемизации. Однако ряд активирующих процедур позволяет снизить степень рацемизации. Наилучшие результаты достигаются использованием ДЦК в смеси с нуклеофильным катализатором, например гидроксибензотриазолом. В этом случае N-ацетиламинокислота ( чаще всего Вос-аминокислота) превращается in situ в активированный эфир, который существенно меньше склонен к образованию азлактона. [32]

После того, как полипептидные цепи отделились от рибосомы, они могут тотчас же приобретать свойственную им вторичную, третичную или четвертичную структуру ( разд. Если рибосома прикреплена к ЭР, то белок поступает в ЭР, откуда затем траспорти-руется к месту назначения. Первая часть растущей полипептидной цепи состоит при этом из сигнальной последовательности аминокислот, которая соответствует определенному рецептору в мембране ЭР, что обеспечивает присоединение рибосомы к ЭР. После того как белок оказывается внутри ЭР, сигнальная последовательность отделяется от него, и белковая молекула свертывается, приобретая свою окончательную форму. [34]
После завершения синтеза получают полистирол с привитыми полипептидными цепями заданного состава. Такой привитой сополимер обрабатывают смесью РзССООН и НВг, что приводит к отщеплению синтезированного полипептида, выделению изобутилена и СО2, а также к регенерации матричного, полимера. Этот процесс синтеза автоматизирован, и современные аминокислотные синтезаторы могут присоединить к растущей полипептидной цепи до 6 аминокислотных звеньев в сутки. [35]
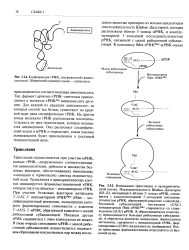
Так, фермент аргинил-т РНК-синтетаза присоединяет к молекуле тРНК аминокислоту аргинин. На другом конце молекулы тРНК расположена последовательность из трех нуклеотидов, которая называется антикодоном. Она распознает специфический кодон в мРНК и определяет, какая именно аминокислота будет присоединена к растущей полипептидной цепи. [37]
С другой стороны, и рибосомы поступают активированные аминокислоты. Через стадию аминоациладенилатов получаются производные аминокислот - аминоацил-т - РНК, к-рые и поступают в рибосомы. Здесь в присутствии м - РНК, ионов магния, гуано-зинтрифосфата происходит распад комплексов аминоацил-т - РНК и включение остатков аминокислот в растущую полипептидную цепь. Наличие в т - РНК участков, комплементарных определенным кодирующим триплетам в м - РНК, обусловливает определенную последовательность расположения остатков аминокислот в синтезируемой полипептидной цепи. [38]
С другой стороны, в рибосомы поступают активированные аминокислоты. Через стадию аминоациладенилатов получаются производные аминокислот - аминоацил-т - РНК, к-рые и поступают в рибосомы. Здесь в присутствии м - РНК, ионов магния, гуано-зинтрифосфата происходит распад комплексов аминоацил-т - РНК и включение остатков аминокислот в растущую полипептидную цепь. Наличие в т - РНК участков, комплементарных определенным кодирующим триплетам в м - РНК, обусловливает определенную последовательность расположения остатков аминокислот в синтезируемой полипептидной цепи. [39]
Было сделано заключение, что трансляция первых кодонов мРНК приводит к синтезу N-концевой сигнальной последовательности, которая определяет присоединение рибосомы к мембране; в течение дальнейшей элонгации эта последовательность отщепляется на мембране, так что конечный продукт оказывается лишенным ее. Вслед за этим было показано, что такая N-концевая последовательность богата гидрофобными аминокислотными остатками; было естественно заключить, что гидрофобный характер сигнальной последовательности растущей полипептидной цепи способствует взаимодействию полипептида с мембраной. [40]
Специфической модификацией полипептидных цепей при синтезе коллагена в клетках соединительных тканей ( фибробластах) животных является гидроксилирование пролиновых и лизиновых остатков. Этот процесс, по крайней мере частично, также котрансля-ционный. Пролил-4 - гидроксилаза, пролил-3 - гидроксилаза и лизил-гидроксилаза, как было показано, - мембраносвязанные ферменты, способные превращать остатки пролина и лизина растущих полипептидных цепей коллагена в остатки 4 - / иранс-гидроксипролина, З - шранс-гидроксипролина и 5-гидроксилизина, соответственно. Дальнейшая модификация полипептидных цепей коллагена включает, в частности, О-гликозилирование оксилизиновых остатков. [41]
Именно в аминоацилированной и формилированной форме ( F-Met - tRNApet) она является инициатором трансляции. Соответственно, первым аминокислотным остатком любой синтезируемой полипептидной цепи в прокариотической рибосоме всегда является формилметионин. В последующем, в ходе элонгации формильный остаток, как правило, отщепляется форми-лазой; метиониновый остаток тоже часто ( но не всегда) отщепляется от растущей полипептидной цепи специальной аминопептидазой. [42]
Во втором случае синтез, включающий 369 химических реакций и 11 931 операцию, был выполнен двумя участниками ( Гатте и Меррифилд) всего за несколько месяцев ( в среднем до шести аминокислотных остатков в день присоединялись к растущей полипептидной цепи - фантастический темп. [43]
Во втором случае синтез, включающий 369 химических реакций и И 931 операцию, был выполнен двумя участниками ( Гатте и Меррифилд) всего за несколько месяцев ( в среднем до шести аминокислотных остатков в день присоединялись к растущей полипептидной цепи - фантастический темп. [44]
Несмотря на большое количество примеров, когда секреторные и внутримембранные белки синтезируются с отщепляемой сигнальной N-концевой последовательностью, это, по-видимому, не есть общее правило. Некоторые белки, как оказалось, тоже синтезируются на мембраносвязанных рибосомах, но без отщепления N-концевой или какой-либо иной сигнальной последовательности. К ним относится такой типичный секреторный белок, как овальбумин, а также мембранные белки цитохром Р-450, эпоксидгидратаза, ретинальный опсин, гликопротеид РЕа вируса Синдбис. В этих случаях растущая полипептидная цепь тоже, по-видимому, имеет сигнальную последовательность, индуцирующую присоединение транслирующей рибосомы к мембране эндоплазматического ретику-лума и ко-трансляционный транспорт белка в мембрану, но без сопутствующего процессинга. Для цитохрома Р-450 показано, что его N-концевая последовательность гидрофобна и напоминает сигнальную, но она сохраняется у зрелого белка. Однако N-концевые последовательности овальбумина и опсина не похожи на обычную сигнальную последовательность. Возможно, что либо сигнальную роль здесь выполняют специальные внутренние гидрофобные участки растущего полипептида, либо не столь гидрофобная N-концевая последовательность тоже в каких-то случаях может служить сигналом для присоединения к мембране. Как и в других случаях, взаимодействие с мембраной возможно лишь в течение трансляции; но не после нее; очевидно, сворачивание завершенной цепи как-то блокирует ( экранирует) сигнальную функцию соответствующего участка. [45]
Страницы: 1 2 3 4