Растущая полипептидная цепь
Cтраница 4
 |
Схема твердофазного синтеза пептидов. [46] |
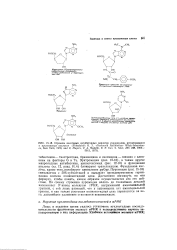
Пептидный синтез в классическом варианте сопряжен со значительными затратами труда и времени. Идея его состоит в закреплении растущей полипептидной цепи на полимерном нерастворимом носителе. При этом значительно упрощаются операции выделения промежуточных продуктов, которые сводятся к экстракции и фильтрованию полимера, полиостью снимается проблема нерастворимости пептидов и создаются предпосылки для автоматизации процесса. Определяющим фактором в твердофазном синтезе является полнота протекания всех химических реакций, которая достигается за счет применения избытка конденсирующего агента и N-защищенной аминокислоты, отделяемых экстракцией. Естественно, выбор защитиык группировок и методов конденсации должен обеспечить полное отсутствие рацемизации. [47]
Эритромицин ( рис. 12 - 10), а также другие макролидные антибиотики, циклогексимид ( рис. 12 - 10) и фузидиевая кислота ( гл. Пуромицин ( рис. 15 - 18) связывается с 505-субчастицей и вызывает преждевременную терми-нацию синтеза полипептидной цепи. Достаточно взглянуть на его формулу, чтобы понять, каким образом осуществляется это его действие. По своему строению пуромицин вплоть до тончайших деталей напоминает З - конец молекулы тРНК, нагруженной аминоацильной группой, с той лишь разницей, что у пуромицина нет аминоацильиой группы, и как только растущая полипептидная цепь переносится на него, дальнейшее удлинение становится невозможным. [49]
С этот фермент синтезируется нормально, тогда как при 37 С процесс трансляции не доходит до конца. Они отметили также, что в клетках, предварительно инкубированных при 25 С, последующий синтез фермента при 37 С мог протекать удовлетворительно. По мнению авторов, это можно было бы объяснить тем, что структурные субъединицы фермента, синтезированные при 25 С, остаются в клетке, а позднее при 37 С действуют как конформационные матрицы. Имея правильную конформацию, они взаимодействуют с растущими полипептидными цепями и придают им надлежащую третичную структуру, что и позволяет довести трансляцию до конца. [50]
Очевидно, что важную роль в ко-трансляционном сворачивании белка может играть образование дисульфидных связей между цистеиновыми остатками. Дисульфидные связи, скрепляющие третичную структуру, особенно распространены у секреторных белков эукариот. Наоборот, внутриклеточные белки чаще характеризуются свободными сульфгидрильными группами цистеиновых остатков. Действительно, условия внеклеточной среды, по сравнению с внутриклеточной, являются более окислительными. Дисульфидные связи, по-видимому, могут завязываться между цистеиновыми остатками растущей полипептидной цепи уже по мере ее прохода через мембрану в межмембранный просвет. Такие связи могут возникать спонтанно при достаточно окислительных условиях среды. Однако, во-первых, скорость спонтанного образования дисульфидных связей в белке, по сравнению со скоростью его синтеза и сворачивания, не велика; во-вторых, в процессе сворачивания всегда существует вероятность образования дисульфидных связей между не теми остатками цистеина, которые должны образовать мостики в законченной свернутой белковой молекуле. [51]
На самом деле, ускоряя как прямую, так и обратную реакцию, фермент катализировал тиол-дисульфид-ный обмен, тем самым приводя не только к быстрому образованию дисульфидных связей, но и к быстрой замене неправильных или промежуточных ( ненативных) связей на правильные в ходе сворачивания белка. Таким образом, фактически катализировался процесс сворачивания дисульфидсшитых белков. Фермент может быть обозначен как дисулъфидизомераза белков. Он очень лабильно ассоциирован с люминальной поверхностью мембраны, обращенной в межмембранный просвет эндоплазматического ретикулума, и не исключено, что он может присутствовать в межмембранном просвете в растворимой форме. Можно думать, что в процессе синтеза белка и выхода растущей полипептидной цепи в межмембранньш просвет дисульфидизомераза катализирует ко-трансляционное образование внутридоменных дисульфидных связей и их перебор в ходе формирования правильной третичной структуры высунувшегося домена. [52]

Этапы элонгации и терминации у про - и эукариот во многом сходны. Рассмотрим процесс более подробно. Когда эта тРНК оказывается на месте, между карбоксильной группой метионина и аминогруппой лейцина с помощью ферментативной активности, присущей большой субъединице, образуется пептидная связь, при этом лейцин остается связанным со своей тРНК, а метионин отсоединяется от инициаторной тРНК, и последняя отделяется от рибосомы. Комплекс метионин-лейцин - тРНК1 - мРНК протягивается через рибосому ( транслокация), так что следующий кодон мРНК может связаться с нагруженной тРНК, несущей соответствующий антикодон. Если третьим кодоном мРНК является UUU, то следующей аминокислотой в растущей полипептидной цепи будет фенил-аланин; его доставит к рибосоме тРНК с антикодоном ААА. [54]
На З - конце такого листа находятся неспаренные основания - последовательность цитозин-цитозин-аденин, на 5 -конце одно неспаренное основание, в основном гуанин. Связывание а-аминокислоты с транспортной РНК осуществляется на З - конце за счет карбоксильной группы аминокислоты. Три другие Цеспаренные специфические основания транспортной РНК образуют триплет ( антикодон), комплементарный кодону матричной РНК. После прикрепления транспортной РНК к информационной РНК ( за счет взаимодействия кодон-антикодон) протекает перенос а-аминокислоты, связанной с транспортной РНК на растущую полипептидную цепь. [55]
Этот триплет оснований взаимодействует с тремя и только с тремя комплементарными основаниями ( ГГУ, например) мРНК, находящимися в контакте с рибосомой. Каждая рибосома передвигается, или скользит, вдоль цепи мРНКтак, что в определенные моменты времени один из ее триплетов оказывается в том месте, куда доставляется аминокислота. Триплет мРНК, определяющий присоединение аминокислоты, таким образом, кодирует ее и называется кодоном. Для триптофана кодоном служит ГГУ. Освободившись от своей тРНК, каждая аминокислота присоединяется ферментативным путем к растущей полипептидной цепи в результате образования пептидной связи между ее аминогруппой и карбоксильным концом растущей цепи. На каждой рибосоме имеется, таким образом, два участка связывания: один - для растущей полипептидной цепи, а другой - для молекул тРНК, приводящих соответствующие аминокислоты. [56]
Этот триплет оснований взаимодействует с тремя и только с тремя комплементарными основаниями ( ГГУ, например) мРНК, находящимися в контакте с рибосомой. Каждая рибосома передвигается, или скользит, вдоль цепи мРНКтак, что в определенные моменты времени один из ее триплетов оказывается в том месте, куда доставляется аминокислота. Триплет мРНК, определяющий присоединение аминокислоты, таким образом, кодирует ее и называется кодоном. Для триптофана кодоном служит ГГУ. Освободившись от своей тРНК, каждая аминокислота присоединяется ферментативным путем к растущей полипептидной цепи в результате образования пептидной связи между ее аминогруппой и карбоксильным концом растущей цепи. На каждой рибосоме имеется, таким образом, два участка связывания: один - для растущей полипептидной цепи, а другой - для молекул тРНК, приводящих соответствующие аминокислоты. [57]
Страницы: 1 2 3 4