Интрона
Cтраница 1
 |
Схема сплайсинга экзонов при образовании зрелой мРНК ( а, границы экзоиов и нитронов ( правило сплайсинга ( б. [1] |
Интроны могут составлять большую часть мозаичного гена. Суммарная нуклеотидная длина интронов может превышать во много раз длину экзонов. Например, в гене проколлагена птиц экзоны составляют лишь восьмую часть общей длины гена, а в гене тиреоглобулина млекопитающих - лишь двадцатую. Экзонами называют не только те районы гена, которые содержат последовательности, кодирующие участки полипептидной цепи, но и районы, транскрипты которых входят в состав зрелых мРНК, но не транслируются. Эти районы соответствуют лидерной области перед инициирующим кодоном ( см. рис. 102) или в нетранслируемой части на 3 -конце мРНК вслед за терминирующим триплетом. [2]
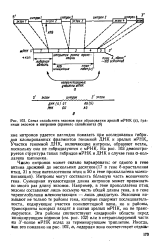 |
Схема сплайсинга экзонов при образовании зрелой мРНК ( а, границы экзоиов и нитронов ( правило сплайсинга ( б. [3] |
Интроны могут составлять большую часть мозаичного гена. Суммарная нуклеотидная длина нитронов может превышать во много раз длину экзонов. Например, в гене проколлагена птиц экзоны составляют лишь восьмую часть общей длины гена, а в гене тиреоглобулина млекопитающих - лишь двадцатую. Экзонами называют не только те районы гена, которые содержат последовательности, кодирующие участки полипептидной цепи, но и районы, транскрипты которых входят в состав зрелых мРНК, но не транслируются. Эти районы соответствуют лидерной области перед инициирующим кодовом ( см. рис. 102) или в нетранслируемой части на 3 -конце мРНК вслед за терминирующим триплетом. [4]
Интроны могут составлять большую часть мозаичного гена. Например, в гене птиц экзоны составляют лишь восьмую часть общей длины гена, а в гене млекопитающих - лишь двадцатую. [5]
 |
Схема сплайсинга экзонов при образовании зрелой мРНК ( а, границы экзоиов и нитронов ( правило сплайсинга ( б. [6] |
Интроны могут составлять большую часть мозаичного гена. Суммарная нуклеотидная длина интронов может превышать во много раз длину экзонов. Например, в гене проколлагена птиц экзоны составляют лишь восьмую часть общей длины гена, а в гене тиреоглобулина млекопитающих - лишь двадцатую. Экзонами называют не только те районы гена, которые содержат последовательности, кодирующие участки полипептидной цепи, но и районы, транскрипты которых входят в состав зрелых мРНК, но не транслируются. Эти районы соответствуют лидерной области перед инициирующим кодоном ( см. рис. 102) или в нетранслируемой части на 3 -конце мРНК вслед за терминирующим триплетом. [7]
 |
Сплайсинг мРНК ( по К. Murray. [8] |
Это так называемые интроны, которые вырезаются из цепи мРНК в процессе ее созревания. Механизм вырезания интронов и сшивания экзонов называется сплайсинг. Механизм точного вырезания интронов и сшивания экзонов достаточно сложен. Его интерпретация стала возможной после того, как было доказано наличие консенсусных последовательностей на границе соединения интронов с экзонами. Основным инструментом сплайсинга являются малые ядерные РНК, обладающие ферментативной активностью. Характерной особенностью рибози-мов является наличие липких концов, комплементарных концам интронов. [9]
Согласно другой гипотезе, интроны разделяют гены на отдельные, способные обмениваться участки ( мини-гены), которые могут рекомбинировать в ходе эволюции вида с образованием новых генов. Какова бы ни была функция нитронов, очевидно, их исследование поможет решить ряд проблем, связанных с транскрипцией генов ( гл. [11]
В генах имеются некодирующие вставки - интроны. [12]
В состав ряда таких генов входят необычно длинные интроны. Возможно, альтернативные пути сплайсинга ( процессинга) длинных первичных транскриптов локуса ВХ приводят к образованию целого набора мРНК, которые транслируются в разных группах клеток с образованием белков, несущих разные функции. Однако детали строения этих генов и особенности регуляции их активности еще мало исследованы. [13]
 |
Сплайсинг мРНК ( по К. Murray. [14] |
Во-первых, в некоторых случаях рибозимы вырезают интроны самостоятельно, без помощи белков сплайсосомы. Следовательно, они обладают каталитической активностью и представляют собой уникальное явление, уточняя и расширяя представление о том, что биологический катализ осуществляется только белками-ферментами. [15]
Страницы: 1 2 3 4