Интрона
Cтраница 4

Поэтому для получения аутентичного фермента кДНК Cu / Zn-SOD человека была встроена в дрожжевой эписомный вектор. Дрожжевые клетки не способны эффективно вырезать интроны, поэтому для кодирования специфичных генных продуктов необходимо использовать соответствующие кДНК или химически синтезированные последовательности. [47]
Одна из возможных функций интронов была обнаружена, когда оказалось, что из одной и той же мРНК в разных клетках могут быть удалены разные интроны. Таким образом, ген может иметь альтернативные интроны и кодировать различные, хотя и сходные белки. Это увеличивает его потенциальное использование. Примером служит ген кальцитонина. Этот ген может продуцировать две разные формы мРНК в зависимости от того, какие интроны были удалены. Одна из них образуется в щитовидной железе и кодирует синтез белка кальцитонина, состоящего из 32 аминокислот. Кальцитонин - это гормон, понижающий уровень кальция в крови. Другая форма мРНК синтезируется в гипоталамусе и кодирует белок, состоящий из 37 аминокислот; этот белок, названный пептидом, связанным с геном кальцитонина, сходен с кальцитонином и обладает сильным сосудорасширяющим действием. Он выделяется также из нервных окончаний в некоторых частях периферической нервной системы. [48]
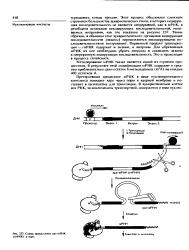 |
Схема процессннга пре-м РНК ( гяРНК в ядре. [49] |
Первичный продукт транскрипции - гяРНК содержит и экзоны, и нитроны. Для образования мРНК из нее необходимо убрать интроны и соединить экзоны в непрерывную кодирующую последовательность. Это и происходит в процессе сплайсинга. [50]
Этот вопрос был выяснен. Оказалось, что эукариотическая РНК-полимераза транскрибирует и экзоны, и интроны, причем точно в той последовательности, в которой они находятся в гене. При этом образуются очень длинные предшественники РНК, которые содержат участки нетранслируемой PHKJ комплементарные нуклеотидным последовательностям интронов ДНК. Такие предшественники мРНК представляют собой значительную часть гяРНК и обнаруживаются только в ядре. [51]
Ферментативная машина сплайсинга, локализованная в ядре, изучена недостаточно, однако ряд важных результатов получен при исследовании превращений предшественников индивидуальных мРНК ( например, р-глобина) в ядерных экстрактах in vitro. Прежде всего происходит разрыв нуклеотидной последовательности на границе экзона 1 и интрона с образованием 5 -конца, представленного фосфор ил ированным гуанозином - каноническим нуклеотидом левого ( 5 -конца) интрона. Следующая ( или сопряженная с предыдущей) реакция заключается во взаимодействии 5 -конца удаляемой последовательности с 2 -гидроксилом аденозина интрона с образованием; 2 - 5 -фосфодиэфирной связи. Образование такой структуры можно также рассматривать как результат реакции трансэтерификации, в результате которой 2 - ОН-группа аденозина атакует фосфодиэфирную связь, замещая З - ОН-группу экзона 1 Теперь экзон 1 представлен отдельной молекулой РНК, удерживаемой вместе с другой молекулой, содержащей лассб-интрон и экзон 2, в едином комплексе с белками, участвующими в реакции. По-видимому, образование лассо является условием для завязывания ковалентной связи З - ОН-группы линейной молекулы экзона 1 с экзоном 2 в районе сайта сплайсинга; эта реакция также представляет собой трансэтерификацию, когда З - ОН-группа экзона 1 атакует фосфодиэфирную связь и замещает З - ОН-группу интрона; последовательность интрона выщепляется из РНК в форме лассо. В результате число фосфодиэфирных связей в составе субстрата и продукта не изменяется. [52]
Ферментативная машина сплайсинга, локализованная в ядре, изучена недостаточно, однако ряд важных результатов получен при исследовании превращений предшественников индивидуальных мРНК ( например, р-глобина) в ядерных экстрактах in vitro. Прежде всего происходит разрыв нуклеотидной последовательности на границе экзона 1 и интрона с образованием 5 -конца, представленного фос-форилированным гуанозином - каноническим нуклеотидом левого ( 5 -конца) интрона. Следующая ( или сопряженная с предыдущей) реакция заключается во взаимодействии 5 -конца удаляемой последовательности с 2 -гидроксилом аденозина интрона с образованием. Образование такой структуры можно также рассматривать как результат реакции трансэтерификации, в результате которой 2 - ОН-группа аденозина атакует фосфодиэфирную связь, замещая З - ОН-группу экзона 1 Теперь экзон 1 представлен отдельной молекулой РНК, удерживаемой вместе с другой молекулой, содержащей лассд-интрон и экзон 2, в едином комплексе с белками, участвующими в реакции. По-видимому, образование лассо является условием для завязывания ковалентной связи З - ОН-группы линейной молекулы экзона 1 с экзоном 2 в районе сайта сплайсинга; эта реакция также представляет собой трансэтерификацию, когда З - ОН-группа экзона 1 атакует фосфодиэфирную связь и замещает З - ОН-группу интрона; последовательность интрона выщепляется из РНК в форме лассо. В результате число фосфодиэфирных связей в составе субстрата и продукта не изменяется. [53]
Ферментативная машина сплайсинга, локализованная в ядре, изучена недостаточно, однако ряд важных результатов получен при исследовании превращений предшественников индивидуальных мРНК ( например, р-глобина) в ядерных экстрактах in vitro. Прежде всего происходит разрыв нуклеотидной последовательности на границе экзона 1 и интрона с образованием 5 -конца, представленного фос-форилированным гуанозином - каноническим нуклеотидом левого ( 5 -конца) интрона. Следующая ( или сопряженная с предыдущей) реакция заключается во взаимодействии 5 -конца удаляемой последовательности с 2 -гидроксилом аденозина интрона с образованием; 2 - 5 -фосфодиэфирной связи. Образование такой структуры можно также рассматривать как результат реакции трансэтерификации, в результате которой 2 - ОН-группа аденозина атакует фосфодиэфирную связь, замещая З - ОН-группу экзона 1 Теперь экзон 1 представлен отдельной молекулой РНК, удерживаемой вместе с другой молекулой, содержащей лассд-интрон и экзон 2, в едином комплексе с белками, участвующими в реакции. По-видимому, образование лассо является условием для завязывания ковалентной связи З - ОН-группы линейной молекулы экзона 1 с экзоном 2 в районе сайта сплайсинга; эта реакция также представляет собой трансэтерификацию, когда З - ОН-группа экзона 1 атакует фосфодиэфирную связь и замещает З - ОН-группу интрона; последовательность интрона выщепляется из РНК в форме лассо. В результате число фосфодиэфирных связей в составе субстрата и продукта не изменяется. [54]
Таким образом, ДНК содержит огромную избыточность: лишь небольшая часть букв используется для формирования белков. По-видимому, такая избыточность нужна для обнаружения и исправления ошибок, т.е. интроны участвуют в системе репарации. Все это напоминает запись на магнитной ленте, когда после информационного файла добавляется контрольная сумма. [55]
РНК) указывает на необходимость соблюдения высочайшей точности процедуры вырезания необходимого гена из ДНК генома соответствующими рестриктазами. В противном случае могут быть получены не структурные транслируемые гены, а интроны или участки экзонов, не кодирующие белок. После того как были разработаны методы искусственного синтеза и сшивки отдельных участков молекулы ДНК, появилась возможность конструирования и создания новых, неизвестных ранее в природе организмов с заранее заданными свойствами. [56]

Самосплайсинг происходит у пре-р РНК простейших ( напр. РНК митохондрий низших грибов и нек-рых др. пре - РНК, у к-рых интроны содержат консервативные последовательности, что обусловливает образование определенных вторичной и третичной структур. [58]
Страницы: 1 2 3 4