Деполяризация - мембрана
Cтраница 1
Искусственная деполяризация мембраны не приводит к активации яйцеклетки; и наоборот, устранение деполяризации мембраны во время оплодотворения не подавляет активацию. Судя по всему, деполяризация нужна лишь для предотвращения полиспермии. [1]
Деполяризация мембран цистерн приводит к высвобождению кальция и началу мышечного сокращения. Кальций связывается с субъединицей С тропонина. Это изменяет конформацию всей молекулы тропонина - субъединица I перестает мешать взаимодействию актина с миозином; изменение конформации субъединицы Т передается на тропомиозин. Далее тропомиозин поворачивается на 20 и открывает закрытые ранее центры в актине для связывания с миозином. Присоединение актина вызывает быстрое освобождение АДФ и Рн из миозина. Поворот головки, связанной с актином, вызывает перемещение тонкой нити относительно миозина. К головке миозина вместо ушедших АДФ и Рн вновь присоединяется АТФ, образуя комплекс М АТФ. Она вновь становится перпендикулярно тонкой нити. В головке миозина, не связанной с актином, происходит гидролиз АТФ. Вновь образуется комплекс АДФ Рн миозин, и все повторяется. После прекращения действия двигательного импульса Са2 с помощью Са2 - зависимой АТФазы переходит в саркоплазматический ре-тикулум. Уход кальция из комплекса тропонина приводит к смещению тропомиозина и закрытию активных центров актина, делая его неспособным взаимодействовать с миозином, - мышца расслабляется. [2]
 |
Электрические токи, возникающие при деполяризации мембраны. [3] |
При деполяризации мембраны возникают токи, замыкающиеся через наружную проводящую среду. [4]
 |
Сравнение потенциала покоя и потенциала действия. [5] |
При деполяризации мембраны возникают токи, замыкающиеся через наружную проводящую среду. Между возбужденным и невозбужденным участками нервного волокна потечет электрический ток, так как у возбужденного участка внутренняя поверхность имеет положительный заряд, а у невозбужденного - отрицательный и между ними возникает разность потенциалов. Этот локальный ток служит раздражителем для невозбужденных участков нервного волокна, непосредственно примыкающих к месту деполяризации. В них также возникает возбуждение, то есть потенциал действия ( или деполяризация), и так далее. По поверхности клетки локальный ток течет от невозбужденного участка к возбужденному; внутри клетки он течет в обратном направлении. Локальный ток, как и любой электрический ток, раздражает соседние невозбужденные участки и вызывает увеличение проницаемости мембраны. Это приводит к возникновению потенциалов действия в соседних участках. В то же время в ранее возбужденном участке происходит восстановительные процессы реполяризации. [6]
Осуществляет блокирование путем деполяризации мембран; не проникает через плацентарный барьер. [7]
Так как для деполяризации мембран в камерах электродиализных опреснительных установок желательно в возможно большей степени турбулизировать поток, нужно, чтобы числа Рейнольдса потока в камерах были больше критических, при которых уже наблюдается квадратичная зависимость между потерей напора в камере и скоростью потока в ней. [8]
В результате притока заряда деполяризация мембраны аксонного холмика постепенно достигает порогового уровня, затем мембрана быстро разряжается, генерируя потенциал действия, н возвращается в первоначальное состояние ( см. ниже); после этого мембранный потенциал вновь достигает порогового уровня, и весь цикл повторяется. Чем больше деполяризующий ток, тем скорее деполяризация достигает пороговой величины н тем выше частота возникновения импульсов. Очень трудно провести точный анализ кодирующего механизма. В нижеследующем чисто качественном описании мы будем употреблять выражение сила синоптической стимуляции или суммарный ПСП, имея в виду тот суммарный ПСП, который имел бы место, если бы генерирование импульсов каким-то образом подавлялось. [9]
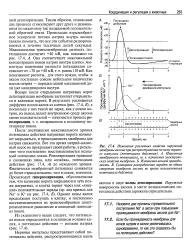
Вскоре после открывания натриевых ворот деполяризация мембраны заставляет открываться и калиевые ворота. [11]
Увеличение внутриклеточного содержания № приводит к деполяризации мембраны, что обусловливает передачу нервного импульса. [12]
Полученные таким образом расходы энергии на деполяризацию мембран представляют собою минимальные значения. [13]
Локальный ( местный) потенциал - потенциал при деполяризации мембраны, который быстро уменьшается по мере удаления от точки возникновения, но не доходит до конца клетки подобно потенциалу действия. Данные потенциалы могут суммироваться. [14]
Величина Q рассчитывается из измерений, например, потенциала деполяризации мембраны, содержащей рецептор. Зависимость обратных величин 1 / Q от 1 / [ L ] может быть использована для графического определения Qmax и Ко так же, как это делается в ферментативной кинетике по методу Лайнувера - Берка. [15]
Страницы: 1 2 3 4