Деполяризация - мембрана
Cтраница 2
Действительно, удалось показать экспериментально, что изгиб волосков приводит к деполяризации мембраны, сопровождаемой возрастанием электрического шума. Одна стереоцилия управляет сорока ионными каналами. Удалось моделировать эти механохимические явления с помощью искусственной бислойной липидной мембраны, в которую встроены ионные каналы. [16]
Взаимодействие рецептора с пахучим веществом приводит к появлению нервного импульса вследствие деполяризации мембраны аксона. Механизм этого процесса пока не ясен. Обонятельный анализатор может воспринимать одну молекулу. [17]
Каждый нервный импульс вызывает в нервно-мышечном синапсе высвобождение достаточного количества ацетилхолина, чтобы деполяризация по-стсинаптической мембраны обусловила сокращение мышечного волокна. [18]
Искусственная деполяризация мембраны не приводит к активации яйцеклетки; и наоборот, устранение деполяризации мембраны во время оплодотворения не подавляет активацию. Судя по всему, деполяризация нужна лишь для предотвращения полиспермии. [19]
Бюлбринг [53] предполагает, что адреналин двояко действует на гладкую мышцу; во-первых, вызывает сокращение в результате непосредственной деполяризации мембраны и, во-вторых, активирует фосфорилазу. Далее Бюлбринг предполагает, что при активации адреналином фосфорилазы выделяется дополнительная энергия для поляризации мембран; таким образом, мембрана стабилизируется и предотвращается спонтанная разрядка. Сетевой эффект должен уменьшить количество сокращающихся клеток, и, следовательно, ткань расслабится. [20]
В принципе быстрые калиевые каналы функционируют таким же образом, как н потенциал-зависимые натриевые: они быстро открываются при деполяризации мембраны н затем полностью инактивируются. Но в отличие от натриевых быстрые калиевые каналы медленнее выходят из состояния инактивации, и для этого требуется возвращение мембранного потенциала к еще более отрицательному уровню. Вероятно, во многих нейронах такого рода градуальное изменение частоты разряда достигается таким же путем. [21]
Порция медиатора, высвобождаемая нервным окончанием в ответ на одиночный нервный импульс, в большинстве случаев оказывается недостаточной для критической деполяризации мембраны гладкомышечной клетки. Критическая деполяризация происходит только при поступлении к нервному окончанию нескольких следующих друг за другом импульсов. Тогда одиночные возбуждающие постсинаптические потенциалы суммируются ( рис. 57) и в момент, когда их сумма достигает пороговой величины, возникает потенциал действия. [22]
Таким образом, согласно мембранной теории при генерации импульса в мембране аксона открывается некий канал, сквозь который ионы Na проникают внутрь аксона, вызывая деполяризацию мембраны. Во время генерации импульса натриевый канал закрывается и открывается калиевый канал. Ионы К выходят наружу, что приводит к восстановлению нормального отрицательного заряда на внутренней стороне мембраны. [23]
Ней-ротоксин; при 10 - 9 - 10 - 7 М ингибирует потенциал действия мембран нервных клеток; блокирует ранний кратковременный поток Na, необходимый для первоначальной деполяризации мембраны нервной клетки. Содержится в некоторых видах рыб ( напр. [24]
Биохимический эффект ацетилхолина заключается в том, что его присоединение к рецептору открывает канал для прохождения ионов Na и К через мембрану клетки, что ведет к деполяризации мембраны. Блокирование действия ацетилхолина чревато серьезными проблемами, вплоть до смертельного исхода. Именно в этом заключается биохимическое действие нейротоксинов. Ниже показаны структуры двух наиболее сильных нейро-токсинов - хистрионикотоксина и хлорида D-тубокурарина. Как и ацетил-холин, молекула D-тубокурарина содержит аммониевые фрагменты. Она блокирует место присоединения ацетилхолина к рецептору, исключает передачу нервного сигнала, предотвращает перенос ионов через мембрану. Создается ситуация, называемая параличом живой системы. [25]
Упрощенная до некоторой степени картина ( рис. 1 - 10) взаимоотношений между изменениями медленных потенциалов и нервными импульсами выглядит следующим образом: внутри нейрона электрохимические процессы ведут к деполяризации мембраны - возрастанию градуальных потенциалов, пока они не становятся нервными импульсами. Там, где соединяются нейроны, или где тонкие, немиелинизированные волокна формируют сплетение и образуют контакты относительно бесструктурным способом, названным эфаптическим ( рис. 1 - 11), или где окончания волокон действительно образуют структурный контакт с другими нейронами посредством синапса ( рис. 1 - 12), нервные импульсы постепенно затухают, то есть превращаются в небольшие медленные потенциалы, не отличимые от спонтанно генерированной локальной градуальной активности ( см. гл. [26]
Взаимосвязь между нарушением ионного гомеостаза и клеточным делением столь очевидна, что А. Г. Маленков формулирует правило, позволяющее предсказать, какие именно химические соединения могут стимулировать деление - это вещества, вызывающие увеличение проницаемости клеточной мембраны для ионов в 2 - 3 раза и деполяризацию мембраны на 50 % и более. Эта зависимость позволила автору сформулировать общебиологический принцип, согласно которому между поведением клетки ( митоз, изменение дифференцировки, переход в режим автономного периодического деления, гибель) и изменениями параметров гомеостаза ( асимптотическое приближение к исходному состоянию, циклические колебания, апериодические изменения) существует однозначное соответствие. Если связь нарушение гомеостаза - гибель тривиальна, то о связи нарушение гомеостаза - развитие этого сказать нельзя. Взаимосвязь клеточного деления с нарушениями гомеостаза установлена и для соматических клеток, например фибробластов или клеток асцитной опухоли Эрлиха. С этим механизмом связано деление клеток регенерирующей печени. [27]
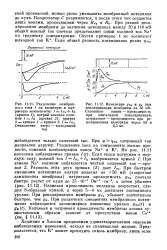
Конденсатор С разряжается, и после этого ток создается лишь ионами, проходящими через Н л и Лк. При резкой деполяризации мембраны до значения потенциала между 20 и 110 мВ общий ионный ток сначала представляет собой входной ток Na по градиенту концентрации. [29]
Положительная обратная связь проявляется в процессе генерации потенциала действия в мембранах. При этом деполяризация мембраны приводит к увеличению проницаемости для ионов натрия, а это ведет к еще большему изменению мембранного потенциала. [30]
Страницы: 1 2 3 4