Киназ
Cтраница 2
 |
Ферменты, активируемые маргаицем. [16] |
В процессах глкжонеогене-за принимают участие пируваткарбоксилаза, катализирующая превращение пирувата в щавелевоуксусную кислоту, а также фосфоенолпируват-варбо кси киназа. У сытых животных в первом из названных ферментов марганец может быть заменен маг - нием и катализируемый им этап глюконеогенеза при дефиците марганца затрагивается, по-видимому, в меньшей степени. Второй фермент из цитозоля печени крыс активируется ионами железа кобальта, кадмия и марганца, но в очищенном виде этот фермент активируется только марганцем. Активность ФЭП-карбок-сикиназы повышается при наличии в инкубационной среде сульфат-аниона [ Bentle L. A., Lardy H. Аналогичный синергизм наблюдается и в действии марганца и сульфат-аниона на глюкозо-6 - фосфатзависимую гликогенсинтазу плаценты человека. [17]
Адреналин стимулирует аденилциклазу, с помощью которой из АТФ образуется циклический АМФ, и затем циклический АМФ активирует киназу, превращающую неактивную фос-форилазу Ь в активную фосфорилазу а ( стр. Активная гликогенсинтетаза L ( так называемая а) превращается в менее ] активную форму D ( так называемая &) под действием киназы, стимулируемой циклическим АМФ. Действие адреналина или глюкагона сводится, таким образом, к подавлению синтеза или ускорению распада гликогена ( стр. [18]
В зараженных клетках значительно возрастает активность киназ, фосфорилирующих ТМФ, и дГМФ, но не наблюдается усиления активности дАМФ - киназы. Большая активность этих ферментов обусловлена их новообразованием, причем новообразованные киназы можно отличить от киназ клеток хозяина, действовавших до заражения. [19]
Перенос АМФ он осуществлять не может, и для ликвидации АМФ, образующегося внутри митохондрий, в межмембранном пространстве находится аденилат киназа, переводящая монофосфат в дифосфат. [20]
Заметим, что в предшествующей работе с участием того же автора [ Lee, Johansson, 1977 ] было показано, что в точном соответствии с характером присоединения фосфатов к аденозину сефарозные сорбенты, несущие на конце шестичленного гидрофобного спейсера АТР, хорошо связывают дегидрогеназы и киназы, зависящие от НАД, а сорбенты, несущие 2 5 - ADP - дегидрогеназы и другие ферменты, зависящие от НАДФ. [21]
Важность катализа определенных стадий фосфотрансферазных реакций обычно очевидна. Киназы и синтетазы уже упомянуты выше. [22]
Ультрацентрифугирование суспензии проводят при 78000 g в течение 100 мин. Супернатант, содержащий киназу фосфорилазы, осторожно декантируют и фильтруют через стекловату. [23]
ФОСФОТРАНСФЕРАЗЫ, то же, что киназы. [24]
Несколько менее подробно обсуждаются и некоторые другие киназы, включая и такие, которые не соответствуют нормальным моделям. [25]
В зараженных клетках значительно возрастает активность киназ, фосфорилирующих ТМФ, и дГМФ, но не наблюдается усиления активности дАМФ - киназы. Большая активность этих ферментов обусловлена их новообразованием, причем новообразованные киназы можно отличить от киназ клеток хозяина, действовавших до заражения. [26]
Фосфорилаза и гликогенсинтаза имеют субъединичное строение. Неактивная ( малоактивная) фосфорилаза b состоит из нефосфори-лированных димеров и под действием фермента киназы фосфорилазы с участием АТФ переходит в фосфорилазу а ( активная форма), которая представлена фосфорилированными тетрамерами. Дефосфори-лирование фермента, т.е. переход формы a - b осуществляется с помощью фермента протеинфосфатазы. Гликогенсинтаза, наоборот, в неактивном состоянии ( D-форма) является фосфорилированным тетрамером. Переход в активную I-форму ( дефосфорилированный тетрамер) осуществляется под действием протеинфосфатазы. [27]
Их совместное действие и определяет в итоге соотношение фосфорилаз а и Ь, а следовательно, и скорость расщепления гликогена с образованием глюкозо-1 - фосфа-та. Естественно задать вопрос: а чем же в свою очередь регулируется активность фосфатазы фосфорилазы и киназы фосфорилазы. [28]
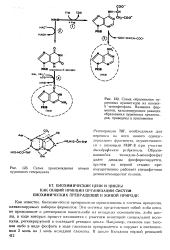 |
Схема образования пу-риновых нуклеотидов из инозин-5 - монофосфата. Названия ферментов, катализирующих реакции образования пуриновых нуклеотидов, приведены в приложении. [29] |
Регенерация TIIF, необходимая для переноса па него нового одиоуг-леродного фрагмента, осуществляется с помощью NADP-II при участии дшидрофолат редуктази. Образовавшийся тпмидин-5 - моиофосфат далее дважды фосфорнлируется, причем на первой стадии пре-ймуществеппо работает специфичная дезокситимидилат киназа. [30]
Страницы: 1 2 3 4