А-химотрипсин
Cтраница 3
В качестве конкретной системы можно исследовать комплекс а-химотрипсина с профлавином, который является ингибитором фермента. [31]
Тщательные исследования [65, 66] показывают, что в а-химотрипсине эта ионная пара стабилизирует активную конформацию фермента на 2 9 ккал / моль. Губер и Воде [94] считают, что ионная пара Asp-194 - Не-16 в трипсине представляет собой нечто вроде скрепки для сложной системы из более чем 20 водородных связей, которыми скреплены сегменты домена активации; в трипсиногене такие связи вообще отсутствуют либо заменяются водородными связями с молекулами воды. [32]
Известно [17], что неконкурентное ингибирование ферментативной активности а-химотрипсина борной кислотой обусловлено взаимодействием ингибитора с имидазольной группой остатка гистидина-57 активного центра фермента. Определить, принимает ли гистидин-57 активного центра фермента участие в связывании н-гексилборной кислоты. [33]
При данной концентрации ДФФ протеиназная и зстеразная активности а-химотрипсина, j - химотрипсина, у-химотрипсина и трипсина в каждом случае ингибируются почти в одинаковой степени. [34]
Парные оси вращения обнаружены в молекулах гемоглобина и а-химотрипсина. Они также могут возникать, например, при компоновке вирусных оболочек из белковых субъединиц. Если эти элементы симметрии совпадают с кристаллографическими осями, они могут быть относительно легко обнаружены методом дифракции рентгеновских лучей. В противном случае их нахождение связано со значительными трудностями. В обоих случаях парные оси вращения обнаруживаются при помощи метода ЭПР значительно легче и проще, чем методом рентгеноструктурного анализа. [35]
Катализируемая химотрипсином элиминация дипептида треониласпараги-новой кислоты от б-химотрипсина дает а-химотрипсин. Тот же продукт может получиться при обратном порядке отщепления дипептидов; в качестве промежуточного продукта в этом случае образуется неохимотрипсино-ген. [36]
На рис. 3 показано пространственное строение молекулы протео-литического фермента а-химотрипсина. [37]
Это позволило авторам предположить, что в реакции гидролиза а-химотрипсином и па-паином сложноэфирных субстратов образуется промежуточное ацилферментное соединение, и гидролиз этого интермедиата является скоростьопределяющей стадией. [38]
Эти наблюдения позволили предположить, что в механизме катализа а-химотрипсином происходит последовательное расщепление сложноэфирной связи с переносом ацильной группы на активный центр фермента, выбросом спиртового продукта и образованием ацилферментного промежуточного соединения. [39]
Важнейшие представители используемых животных протеи-наз - пепсин, трипсин, а-химотрипсин и сычужный фермент, иногда называемый реннином или химозином. Пепсин образуется из пепсиногена слизистой оболочки желудка, рН - оптимум его действия на белки - 1 7 - 2 2; он расщепляет их, как и пептиды, по связям, которые образованы с участием аминной группы тирозина или фенил-аланина. Трипсин образуется из трипсиногена поджелудочной железы, его действие наиболее интенсивно при рН 7 6 - 9 0; специфичность четко выражена: он гидролизует связи, сформированные с участием СООН-группы положительно заряженных аминокислот, аргинина и лизина. Химотрипсин близок по ряду свойств трипсину, а специфичность его подобна пепсину с тем отличием, что он разрывает связи, в которые ароматические аминокислоты входят не аминными, а карбоксильными группами. Реннин образуется из прореннина в сычуге молодых жвачных животных. Это - протеолитический фермент, специально настроенный на створаживание молока. [40]
В последнее время работами Хесса с сотрудниками [5-7] на примере а-химотрипсина был развит новый метод изучения кинетики начальных стадий ферментативных реакций, получивший название метода вытеснения профлавина. Метод основан на том факте, что краситель профлавин ( 3 6-диаминоакридин) при связывании с а-химотрипсином в водном растворе изменяет свой спектр поглощения в ультрафиолетовой области. Величина разностного спектра поглощения, имеющего максимальное значение при длине волны 465 нм, пропорциональна концентрации комплекса фермент-профлавин. Введение в систему фермент-профлавин субстрата, конкурирующего с красителем за связывание на активном центре а-химотрипсина, приводит к двум последовательным процессам вытеснения профлавина. Первый, очень быстрый процесс, заключается в обратимом вытеснении красителя из комплекса его с ферментом за счет образования нековалентного фермент-субстратного комплекса. Второй процесс, времена прохождения которого лежат обычно в пределах разрешения установок типа остановленной струи, вызван химическим взаимодействием субстрата с ферментом ( например, образованием ацилферментного промежуточного соединения), что приводит к дополнительному уменьшению концентрации комплекса фермент-профлавин. Изучение кинетики второго процесса при различных концентрациях субстрата в дополнение к изучению кинетики ферментативной реакции в стационарном режиме позволяет сделать заключения о стадийности изучаемой реакции, а также найти значения констант скоростей промежуточных стадий ферментативной реакции. [41]
При изучении методом температурного скачка кинетики связывания профлавина с активным центром а-химотрипсина было обнаружено, что кинетическая кривая является суммой двух экспоненциальных зависимостей. [42]
Асимметрическое ацетилирование () - бутанола-2 гари применении / г-нитро-фенилацетата и а-химотрипсина. [43]
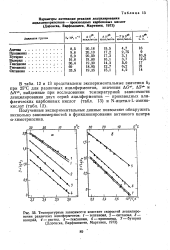
Полученные экспериментальные данные позволяют обнаружить несколько закономерностей в функционировании активного центра а-химотрипсина. [45]
Страницы: 1 2 3 4