Кофейная кислота
Cтраница 2
Существуют два изомерных монометиловых эфира кофейной кислоты: феруловая кислота ( из смолы Ferula и смолы черных сосен) и гесперетиновая кислота ( например, из гесперетина, стр. [16]
Принимая во внимание сравнительно высокую токсичность кофейной кислоты, можно предположить, что это соединение является одним из факторов устойчивости растений к патогенам. Токсичное действие этого фенола, по-видимому, связано с нарушением нормального хода дыхания как тканей растения, так и патогена. Проявление полученной хроматограммы AgNCb и анилинфталатом показало, что в мицелии, выращенном на обычной среде, содержится 5 хорошо разделившихся Сахаров, из которых были идентифицированы ра-финоза, мальтоза, сахароза и глюкоза. В варианте с кофейной кислотой была обнаружена иная картина. Реакция с анилинфталатом свидетельствовала о том, что это пятно представляет собой скопление неразделившихся Сахаров. Поскольку сахара являются одним из основных субстратов дыхания, такое накопление их в мицелии, выращенном на среде с кофейной кислотой, свидетельствовало о резком нарушении нормального хода дыхательного процесса у гриба. [17]
Кортикостерон, производное 401 Коупа перегруппировка 533 Кофейная кислота 177 Коха - Хаафа реакция 90 Крама правило 520 Краун-эфиры 122, 134, 412 ел. [18]
В природе находятся два простых монометиловых эфира кофейной кислоты. Один из них - - окси-3 - метоксикоричная кислота, или феруловая кислота, была выделена из смолы asa foetida, извлеченной из растения ferula семейства зонтичных. Изоферуловая кислота, или гесперетиновая кислота ( Ь - метокси - З - окси-корич-ная кислота), получается в результате расщепления глюкозида гесперетина, выделенного из незрелых апельсинов. [19]
 |
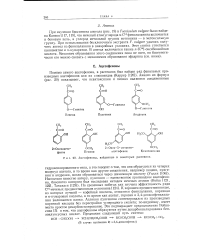
Механизм образования производных коричной кислоты. [20] |
Оказалось также, что фенилаланин является предшественником кофейной кислоты в растениях табака / 23, 24 /, где последняя находится в форме хлорогеновой кислоты. [21]
 |
Изменение содержания хлорогеновой кислоты в периферийном слое клубней картофеля в период формирования клубней в зависимости от интенсивности выпадения осадков. [22] |
Более того, в период покоя содержание кофейной кислоты в поражаемых сортах было заметно выше, чем в устойчивых. Вместе с тем эти сорта значительно различались по устойчивости к фитофторе. [23]
Сложные эфиры п-кумаровой кислоты отличаются от эфиров кофейной кислоты тем, что они не флуоресцируют в ультрафиолетовом свете в отсутствие паров аммиака. Ультрафиолетовые спектры этих сложных эфиров похожи на спектры свободных кислот; длинноволновые полосы в этаноле наблюдаются при 312 - 314 и 330 - 335 ммк соответственно и сдвиги в щелочах - от 50 до 65 ммк. NaOH) при комнатной температуре, эстеразой и 3-глюкозидазой ( только сложные эфиры глюкозы); продукты гидролиза можно идентифицировать хроматографически. [24]
Были обнаружены ферменты, которые катализируют 0-метилиро-вание кофейной кислоты; вероятно, очень близкие к ним ферменты действуют на антоцианы. [25]
 |
Ацетофеноны, найденные в некоторых растениях. [26] |
К хорошим предшественникам, из которых лучший - кофейная кислота, относятся фенилаланин, коричная и n - кумаровая кислоты, в то время как ацетат, тирозин и 3 4-диоксифенилала-нин включаются плохо. Агликон пунгенина синтезировался из производных коричной кислоты без перестройки углеродного скелета; по-видимому, имеет место простое декарбоксилирование. Это подтверждает предположение Робинсона [5] о том, что ацетофеноны образуются путем декарбоксилирования бен-зоилуксусных кислот. [27]
Показано, что вещество, считавшееся сложным производным кофейной кислоты, имеет другую структуру. [28]
Вместе с тем если гидроксилирование предшествует конденсации, то кофейная кислота должна быть наиболее эффективным предшественником кверцитина. Нужно сказать, однако, что если кофейная кислота принимает участие в конденсации в виде производного КоА, то эффективность ее будет ограничиваться скоростью, с которой она активируется. [29]
Следует отметить, что характер изменения величины отношения содержания кофейной кислоты к содержанию хлорогеновой кислоты в некрозе и пораженной ткани в ходе хранения ( рис. 3) очень близок к характеру изменений этой величины в покровных тканях изучаемых сортов. [30]
Страницы: 1 2 3 4