Кооперативное взаимодействие
Cтраница 1
Кооперативные взаимодействия часто проявляются в хорошо известных сигмоидальных кривых, связывающих ферментативную активность с концентрацией субстрата или активатора. Это означает также, что связывание с отдельными центрами дает больше, чем простой аддитивный эффект, и указывает на то, что должно происходить некоторое взаимодействие связывающих центров между собой. [1]
Кооперативное взаимодействие в полимерных цепях, оказывающее существенное влияние на анизотропный рост макромолекулы, может быть обусловлено наличием водородных связей типа тех, которые встречаются у пептидных групп СО - NH. Благодаря этим связям вращение в мономерном кристалле также отличается кооперативностью, напоминающей вращение шестеренок, находящихся в зацеплении. [2]
Кооперативное взаимодействие каталитически активных групп. Многоцентровые катализаторы гораздо более эффективны, чем обычные. Именно отим объясняют высокую эффективность ферментативных реакций, в частности катализируемых протеолитич. При переходе от бимолекулярного к полифункциопаль-пому катализу энтальпия активации переходного состояния сольволиза снижается. [3]
Кооперативное взаимодействие каталитически активных групп. Многоцентровые катализаторы гораздо более эффективны, чем обычные. Именно этим объясняют высокую эффективность ферментативных реакций, в частности катализируемых протеолитич. При переходе от бимолекулярного к полифункциональному катализу энтальпия активации переходного состояния сольволиза снижается. [4]
Кооперативные взаимодействия типа олигомер - полимер и полимер - полимер играют важную роль в стабилизации различных биологических структур. [5]
Хорошо видно кооперативное взаимодействие процессов сдвига и отрыва, формирующее в конечном итоге строение поверхности макротрещины. Следовательно, при анализе самоподобия изломов и связи шероховатости поверхности с фрактальной размерностью структуры излома ( и конечном итоге и с ее свойствами) требуется учет этого взаимодействия. [6]
Однако отсутствие кооперативного взаимодействия субьединиц в окислительно-восстановительной реакции при рН 6 не может быть обусловлено только различной ориентацией порфирина относительно ближайшего белкового окружения гемовых групп. Кооперативное взаимодействие является следствием передачи структурных изменений, берущих начало с изменения ионного радиуса железа и завершающихся при передаче структурных изменений через поверхностные области субъединиц. Как указывалось выше, ответственными за передачу эффекта изменения ионного радиуса оказываются несвязывающие взаимодействия каркаса порфирина с белковым окружением. Увеличение рН не только вызывает увеличение доли низкоспиновых окисленных производных предположительно с копланарной конфигурацией железопорфирина, но также может приводить к ионизации боковых цепей аминокислот, переводя их в форму, благоприятную для несвязывающих взаимодействий с порфириновым кольцом низкоспинового производного. Тем не менее эти малые изменения необходимы для полного проявления кооперативного взаимодействия субъединиц гемоглобина. [7]
Второй важный случай кооперативных взаимодействий проявляется в системах, построенных из нескольких субъединиц, содержащих однотипные центры связывания лиганда L. Важной особенностью этой структуры является тот факт, что присоединение 02 к одной из субъединиц вызывает конформацион-ное изменение не только в ней самой, но и в контактирующих с ней субъединицах, причем в результате этих изменений повышается их сродство к кислороду. Важно, что при этом зависимость становится значительно более благоприятной для выполнения гемоглобином его основной биологической функции - переноса кислорода от легких к тканям живого организма. [8]
При пороговых перегрузках возникает кооперативное взаимодействие реакции пульпозного ядра и внешней нагрузки, приводящее к раскрытию питательных каналов. Это обеспечивает спонтанный высокоскоростной ток пульпозной жидкости в тра-бекулярное пространство, сопровождаемый генезисом диссипативной структуры внутреннего норового скелета позвонка. При этом образующаяся диссипативная структура способна обеспечить работоспособность позвонка при значительных избыточных давлениях. Управляющим синер-гетическим параметром в данном процессе служит разность давлений, определяющая скорость течения пульпозной жидкости. Переход к неустойчивому току жидкости соответствует точке бифуркации, контролируемой числом Рейнольдса. [9]
Принципиальная возможность осуществления физического механизма кооперативного взаимодействия доказана экспериментально. [10]
Сложные системы формируются в результате кооперативных взаимодействий, приводящих к синергетическим ( неаддитивным, нелинейным, резонансным) эффектам. [11]
Все эти полиэлектролиты способны вступать в кооперативное взаимодействие с комплементарными структурами биосистем и образовывать интерполимерные комплексы за счет электростатических и гидрофобных связей. [12]
Уменьшение концентрации деформирующих ионов, ослабляющее кооперативные взаимодействия, оказывает точно такое же влияние, как и повышение температуры. [13]
Разрыв гликозидной связи осуществляется в результате кооперативного взаимодействия двух карбоксильных групп ( в случае лизоцима - это карбоксильные группы глутаминовой кислоты 35 и аспарагиновой кислоты 52 [1]), расположенных в непосредственной близости от расщепляемой связи по двум ее сторонам. [14]
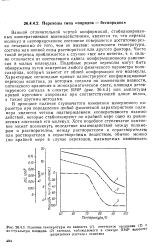 |
Влияние температуры на вязкость ( /, оптическое вращение 3 я интегральную площадь ( 3 сигнала, наблюдаемого в спектре ЯМР высоко. разрешения раствора ксантана. [15] |
Страницы: 1 2 3 4